Функции и строение оболочки клетки
3. Строение клетки. Клеточные органоиды
Ядрышко представляет собой плотное округлое тело внутри ядра. Обычно в ядре клетки бывает от одного до семи ядрышек. Они хорошо видны между делениями клетки, а во время деления — разрушаются.Функция ядрышек — синтез РНК и белков, из которых формируются особые органоиды — рибосомы.
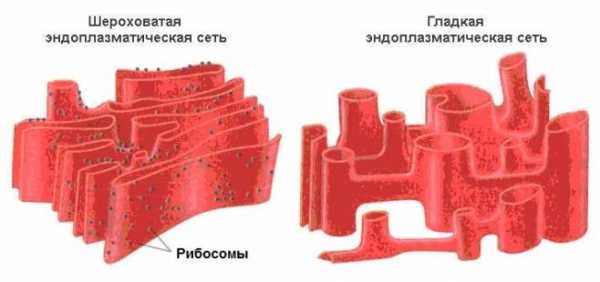
Рибосомы участвуют в биосинтезе белка. В цитоплазме рибосомы чаще всего расположены на шероховатой эндоплазматической сети. Реже они свободно взвешены в цитоплазме клетки.
Эндоплазматическая сеть (ЭПС) участвует в синтезе белков клетки и транспортировке веществ внутри клетки.
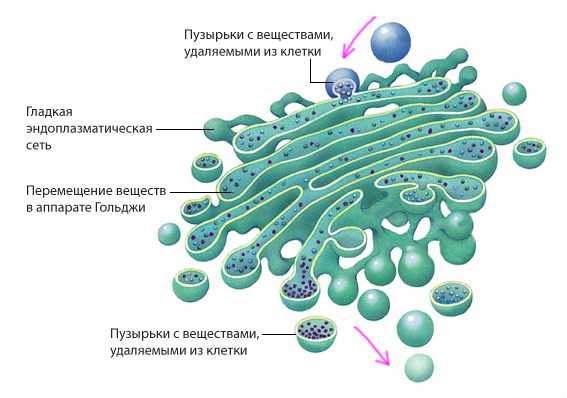
Значительная часть синтезируемых клеткой веществ (белков, жиров, углеводов) не расходуется сразу, а по каналам ЭПС поступает для хранения в особые полости, уложенные своеобразными стопками, «цистернами», и отграниченные от цитоплазмы мембраной. Эти полости получили название аппарат (комплекс) Гольджи. Чаще всего цистерны аппарата Гольджи расположены вблизи от ядра клетки.
Аппарат Гольджи принимает участие в преобразовании белков клетки и синтезирует лизосомы — пищеварительные органеллы клетки.
Лизосомы представляют собой пищеварительные ферменты, «упаковываются» в мембранные пузырьки, отпочковываются и разносятся по цитоплазме.
В комплексе Гольджи также накапливаются вещества, которые клетка синтезирует для нужд всего организма и которые выводятся из клетки наружу.
Митохондрии — энергетические органоиды клеток. Они преобразуют питательные вещества в энергию (АТФ), участвуют в дыхании клетки.
Митохондрии покрыты двумя мембранами: наружная мембрана гладкая, а внутренняя имеет многочисленные складки и выступы — кристы.
В мембрану крист встроены ферменты, синтезирующие за счёт энергии питательных веществ, поглощённых клеткой, молекулы аденозинтрифосфата (АТФ).
АТФ — это универсальный источник энергии для всех процессов, происходящих в клетке.
Количество митохондрий в клетках различных живых существ и тканей неодинаково.
Например, в сперматозоидах может быть всего одна митохондрия. Зато в клетках тканей, где велики энергетические затраты (в клетках летательных мышц у птиц, в клетках печени), этих органоидов бывает до нескольких тысяч.
Митохондрии имеют собственную ДНК и могут самостоятельно размножаться (перед делением клетки число митохондрий в ней возрастает так, чтобы их хватило на две клетки).
Митохондрии содержатся во всех эукариотических клетках, а вот в прокариотических клетках их нет. Этот факт, а также наличие в митохондриях ДНК позволило учёным выдвинуть гипотезу о том, что предки митохондрий когда-то были свободноживущими существами, напоминающими бактерии. Со временем они поселились в клетках других организмов, возможно, паразитируя в них. А затем за многие миллионы лет превратились в важнейшие органоиды, без которых ни одна эукариотическая клетка не может существовать.
Плазматическая мембрана
Строение клетки - все компоненты клеток с определениями
Вы можете почитать про клеточную теорию, которую мы изучили на предыдущем занятии, сегодня мы рассмотрим строение клетки. Строение клетки — это продолжение биологии в рамках понятий клеточной теории.
В этом уроке мы поговорим об обязательной структуре всего живого – клетке. Клетка — структурно-функциональная элементарная единица строения и жизнедеятельности всех организмов (кроме вирусов, о которых нередко говорят, как о неклеточных формах жизни), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению (животные, растения и грибы), либо является одноклеточным организмом (многие простейшие и бактерии). Раздел биологии, занимающийся изучением строения и жизнедеятельности клеток, получил название цитологии. В последнее время принято также говорить о биологии клетки, или клеточной биологии.
Строение клеток
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
- прокариоты (доядерные) — более простые по строению и возникли в процессе эволюции раньше;
- эукариоты (ядерные) — более сложные, возникли позже. Клетки, составляющие тело человека, в основном, являются эукариотическими.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам.
Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органоиды и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждый из органоидов клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
Прокариоты (от лат. Pro — перед, до и греч. Κάρῠον — ядро, орех) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
Эукариоты (эвкариоты) (от греч. Ευ — хорошо, полностью и κάρῠον — ядро, орех) — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.
Строение прокариотической клетки
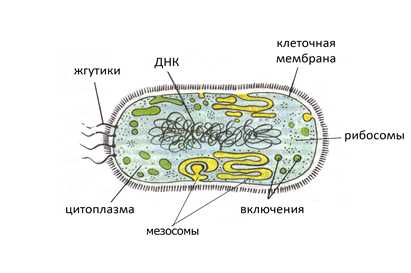
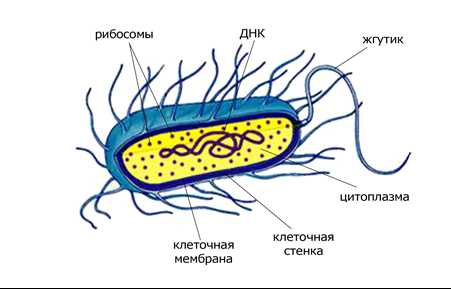
Рисунок 1. Прокариотическая клетка бактерий
Клетки двух основных групп прокариот — бактерий и архей — похожи по структуре: характерными их признаками являются отсутствие ядра и мембранных органелл.
Основные компоненты прокариотической клетки
Основными компонентами прокариотической клетки являются:
- Клеточная стенка, которая окружает клетку извне, защищает её, придаёт устойчивую форму, предотвращающую от осмотического разрушения. У бактерий клеточная стенка состоит из муреина, построенного из длинных полисахаридных цепей, соединенных между собой короткими пептидными перемычками. Клеточная стенка архей не содержит муреина, а построена в основном из разнообразных белков и полисахаридов.
- Жгутики — органеллы движения некоторых бактерий. Бактериальный жгутик построен значительно проще эукариотического, и он в 10 раз тоньше, внешне не покрыт плазматической мембраной и состоит из одинаковых молекул белков, которые образуют цилиндр. В мембране жгутик закреплен при помощи базального тела.
- Плазматическая и внутренние мембраны. Общий принцип устройства клеточных мембран не отличается от эукариот, однако химическом составе мембраны есть немало различий, в частности, в мембранах прокариот отсутствуют молекулы холестерина и некоторых липидов, присущих мембранам эукариот. Большинство прокариотических клеток (в отличие от эукариотических) не имеют внутренних мембран, которые разделяют цитоплазму на отделы (компартменты). Только у некоторых фотосинтетических и аэробных бактерий плазмалемма образует вгибание внутрь клетки, что выполняет соответствующие метаболические функции.
- Нуклеоид — не ограниченный мембранами участок цитоплазмы, в котором расположена кольцевая молекула ДНК — «бактериальная хромосома», где хранится весь генетический материал клетки.
- Плазмиды — небольшие дополнительные кольцевые молекулы ДНК, несущие обычно всего несколько генов. Плазмиды, в отличие от бактериальной хромосомы, не являются обязательным компонентом клетки. Обычно они придают бактерии определенные полезные для неё свойства, такие как устойчивость к антибиотикам, способность усваивать из среды определенные энергетические субстраты, способность инициировать половой процесс и тд.
- Рибосомы прокариот, как и у всех других живых организмов, отвечают за осуществление процесса трансляции (одного из этапов биосинтеза белка). Однако бактериальные рибосомы несколько меньше, чем эукариотические и имеют другой состав белков и РНК. Из-за этого бактерии, в отличие от эукариот, чувствительны к таким антибиотикам, как эритромицин и тетрациклин, которые избирательно действуют на прокариотические рибосомы.
- Споры (эндоспоры) — окруженные плотной оболочкой структуры, содержащие ДНК бактерии и обеспечивающее выживание в неблагоприятных условиях. К образованию спор способны лишь некоторые виды прокариот, например в частности возбудитель столбняка, возбудитель ботулизма и возбудитель сибирской язвы. Для образования эндоспоры клетка реплицирует свою ДНК и окружает копию плотной оболочкой, из созданной структуры удаляется избыток воды, и в ней замедляется метаболизм. Споры бактерий могут выдерживать довольно жесткие условия среды, такие как длительное высушивание, кипячение, коротковолновое облучение.
Сравнительная характеристика клеток эукариот и прокариот
Вы можете увидеть сравнение по признакам прокариот и эукариот в таблице.
| Признак | Прокариоты | Эукариоты |
| Размеры клеток | Средний диаметр 0,5 —10 мкм | Средний диаметр 10 — 100 мкм |
| Организация генетического материала | ||
| Форма, количество и расположение молекул ДНК | Обычно имеется одна кольцевая молекула ДНК, размещенная в цитоплазме | Обычно есть несколько линейных молекул ДНК — хромосом, локализованных в ядре. В интерфазном ядре (вне деления) хромосомы представляют собой хроматин: ДНК компактизируется в комплексе с белками |
| Деление | ||
| Тип деления | Простое бинарное деление. Веретено деления не образуется | Мейоз или митоз |
| Органеллы | ||
| Наличие мембранных органелл | Окруженные мембранами органеллы отсутствуют, иногда плазмалемма образует выпячивание внутрь клетки | Имеется большое количество одномембранных и двумембранных органелл |
Строение эукариотической клетки
Строение эукариотической клетки смотрите на рисунке.
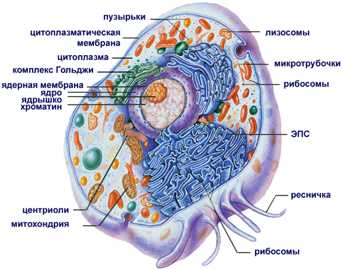
Рисунок 2. Строение эукариотической клетки
Поверхностный комплекс клетки
Плазматическая мембрана называется также плазмалеммой, наружной клеточной мембраной. Это биологическая мембрана, толщиной около 10 нанометров. Обеспечивает в первую очередь разграничительную функцию по отношению к внешней для клетки среде. Кроме этого она выполняет транспортную функцию.
Поверхностый аппарат животных клеток дополнительно включает гликокаликс. Гликокаликс представляет собой «заякоренные» в плазмалемме молекулы углеводов. Гликокаликс выполняет рецепторную и маркерную функции.
У большинства грибов и растений есть клеточная стенка — жёсткая оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции.
Рисунок 3. Клеточная мембрана.
Мембрана клетки
- Барьер толщиной около 8 нм, отделяет живую клетку от окружающей ее среды
- Фосфолипиды и белки являются основными макромолекулами в мембранах
- Составляющие являются амфипатическими молекулами
- Имеются гидрофобные и гидрофильные области
- Избирательно проницаема, что позволяет некоторым веществам проходить легче, чем другим.
Цитоплазма
Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено.
Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами», и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
Эндоплазматический ретикулум
В эукариотической клетке существует система переходящих друг в друга мембранных отсеков (трубок и цистерн), которая называется эндоплазматическим ретикулумом (или эндоплазматическая сеть, ЭПР или ЭПС). Ту часть ЭПР, к мембранам которого прикреплены рибосомы, относят к шероховатому (гранулярному, грубому) эндоплазматическому ретикулуму, на его мембранах происходит синтез белков. Те компартменты, на стенках которых нет рибосом, относят к гладкому ЭПР, принимающему участие в синтезе липидов. Внутренние пространства гладкого и гранулярного ЭПР не изолированы, а переходят друг в друга и сообщаются с просветом ядерной оболочки.
Рисунок 4. Эндоплазмический ретикулум
Гранулярный ЭПР
- Расположены в плоских мешках
- Рибосомы на поверхности придают ей грубый вид
- Некоторые полипептидные цепи входят в грубый ЭПР и модифицированы
- Клетки, которые специализируются на секретировании белков, имеют много грубых ЭПР
Гладкий ЭПР
- Серия взаимосвязанных трубочек
- На поверхности нет рибосом
- Липиды собраны внутри канальцев
- Гладкая ЭПР печени инактивирует отходы, лекарства
- Саркоплазматическая сеть мышц является специализированной формой, которая хранит кальций
Функции ЭПР
Гладкий ЭПР
- Синтезирует липиды
- Метаболизирует углеводы
- Детоксифицирует лекарства и яды
- Накапливает ионы кальция
Гранулированный ЭПР
- Имеет связанные рибосомы
- Распределяет транспортные пузырьки, белки, окруженные мембранами
- Является мембранным заводом для клетки
- Аппарат Гольджи
Аппарат Гольджи
Аппарат Гольджи представляет собой стопку плоских мембранных цистерн, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. То есть это органоид, который упаковывает синтезированные в клетке вещества и побочные продукты для дальнейшей секреции или расщепления.

Рисунок 5. Аппарат Гольджи
Функции аппарата Гольджи
- Модифицирует продукты клетки.
- Производит определенные макромолекулы.
- Сортирует и упаковывает материалы в транспортные пузырьки.
Пузырьки
Небольшие мембранно-замкнутые мешковидные пузырьки образуются в большом количестве во множестве типов, как сами по себе, так и в почках. Есть много типов, но два основных: лизосомы и пероксисомы.
Лизосомы
Лизосомы, которые исходят из органов Гольджи, принимают участие во внутриклеточном пищеварении. Они содержат мощные ферменты, которые могут расщеплять углеводы , белки, нуклеиновые кислоты и липиды. Везикулы внутри лейкоцитов или амеб доставляют лизосомы к поглощенным бактериям, клеточным частям и другому мусору. Ферменты работают лучше всего в кислой среде внутри лизосомы.
Лизосомы разрушают изношенные части клеток или молекулы, чтобы их можно было использовать для создания новых клеточных структур. Некоторые типы клеток могут поглощать другие клетки путем фагоцитоза; это формирует пищевую вакуоль. Лизосома сливается с пищей вакуолизирует и переваривает молекулы. Лизосомы также используют ферменты для рециркуляции собственных органелл и макромолекул клетки, процесс, называемый аутофагией.
Пероксисомы
У растений и животных пузырьки, называемые пероксисомами, образуют и делятся сами по себе, поэтому они не являются частью эндомембранной системы.
Пероксисомы содержат ферменты, которые переваривают жирные кислоты и аминокислоты. Они также расщепляют перекись водорода, токсичный побочный продукт метаболизма жирных кислот.
Ферменты пероксисом превращают перекись водорода в воду и кислород или используют ее в реакциях, которые расщепляют алкоголь и другие токсины.
Ядро
Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки), после чего они выходят в цитоплазму.
Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками. Оболочка ядра двумембранная, сливается с шероховатым ЭПР. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой.
Рисунок 6. Ядро клетки.
Вакуоль
Вакуоль — одномембранный органоид, содержащийся в некоторых эукариотических клетках и выполняющий различные функции (секреция, экскреция и хранение запасных веществ, аутофагия, автолиз и др.). Вакуоли развиваются из мембранных пузырьков — провакуолей. Провакуоли являются производными эндоплазматического ретикулума и комплекса Гольджи, они сливаются и образуют вакуоли.
Рисунок 7. Вакуоль.
Вакуоли и их содержимое рассматриваются как обособленный от цитоплазмы компартмент. Различают пищеварительные и сократительные (пульсирующие) вакуоли, регулирующие осмотическое давление и служащие для выведения из организма продуктов распада. Вакуоли особенно хорошо заметны в клетках растений: во многих зрелых клетках растений они составляют более половины объёма клетки, при этом они могут сливаться в одну гигантскую вакуоль. Одна из важных функций растительных вакуолей — накопление ионов и поддержание тургора (тургорного давления). Вакуоль — это место запаса воды.
Мембрана, в которую заключена вакуоль, называется тонопласт, а содержимое вакуоли — клеточный сок. Клеточный сок состоит из воды и растворенных в ней веществ.
Цитоскелет
К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные филаменты. Микротрубочки принимают участие в транспорте органелл, входят в состав жгутиков, из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, по-видимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка.
Центриоли
Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет, за исключением низших водорослей). Центриоль представляет собой цилиндр, боковая поверхность которого образована микротрубочками.
Вокруг центриолей находится так называемый центр организации цитоскелета, район в котором группируются минус концы микротрубочек клетки.
Рисунок 8. Центриоли.
Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путём синтеза новой структуры, перпендикулярной существующей.
Митохондрии
Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. Дыхание (поглощение кислорода и выделение углекислого газа) происходит также за счёт энзиматических систем митохондрий.
Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами, наружной и внутренней, между которыми располагается межмембранное пространство. Внутренняя мембрана митохондрии образует складки, так называемые кристы. В матриксе содержатся различные ферменты, принимающие участие в дыхании и синтезе АТФ. Центральное значение для синтеза АТФ имеет водородный потенциал внутренней мембраны митохондрии.
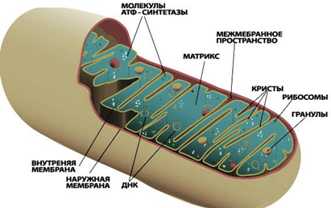
Рисунок 9. Митохондрии.
Митохондрии имеют свой собственный ДНК-геном и прокариотические рибосомы, что, безусловно, указывает на симбиотическое происхождение этих органелл. В ДНК митохондрий закодированы совсем не все митохондриальные белки, большая часть генов митохондриальных белков находятся в ядерном геноме, а соответствующие им продукты синтезируются в цитоплазме, а затем транспортируются в митохондрии. Геномы митохондрий отличаются по размерам: например геном человеческих митохондрий содержит всего 13 генов.
Пластиды
Пластиды (от др.-греч. Πλαστόс — вылепленный) — полуавтономные органеллы высших растений, водорослей и некоторых фотосинтезирующих простейших. Пластиды имеют от двух до четырёх мембран, собственный геном и белоксинтезирующий аппарат.
Согласно симбиогенетической теории пластиды, как и митохондрии, произошли в результате «захвата» древней цианобактерии предшественником эукариотической «хозяйской» клетки. При этом внешняя мембрана пластид соответствует плазматической мембране хозяйской клетки, межмембранное пространство — внешней среде, внутренняя мембрана пластид — мембране цианобактерии, а строма пластид — цитоплазме цианобактерии. Наличие трёх (эвгленовые и динофлагелляты) или четырёх (золотистые, бурые, жёлто-зелёные, диатомовые водоросли) мембран считается результатом двух- и трёхкратного эндосимбиоза соответственно.
Хлоропласты (от греч. Χλωρός — «зелёный») — зелёные пластиды, которые встречаются в клетках фотосинтезирующих эукариот. С их помощью происходит фотосинтез. Хлоропласты содержат хлорофилл.
В одной клетке листа может находиться 15—20 и более хлоропластов, а у некоторых водорослей — лишь 1 -2 гигантских хлоропласта (хроматофора) различной формы.
Хлоропласты ограничены двумя мембранами — наружной и внутренней. Наружная мембрана отграничивает жидкую внутреннюю гомогенную среду хлоропласта — строму (матрикс) В строме содержатся белки, липиды, ДНК (кольцевая молекула) , РНК, рибосомы и запасные вещества (липиды, крахмальные и белковые зерна), а также ферменты, участвующие в фиксации углекислого газа.
Внутренняя мембрана хлоропласта образует впячивания внутрь стромы — тилакоиды, которые имеют форму уплощенных мешочков (цистерн) . Несколько таких тилакоидов, лежащих друг над другом, образуют грану, и в этом случае они называются тилакоидами граны. Именно в мембранах тилакоидов локализованы светочувствительные пигменты, а также переносчики электронов и протонов, которые участвуют в поглощении и преобразовании энергии света.
Рисунок 10. Хлоропласты.
Межклеточные контакты
У высших животных и растений клетки объединены в ткани и органы, в составе которых они взаимодействуют между собой, в частности, благодаря прямым физическим контактам. В растительных тканях отдельные клетки соединяются между собой с помощью плазмодесм, а животные образуют различные типы клеточных контактов, в основном десмосомы.
Плазмодесмы растений — это тонкие цитоплазматические каналы, которые проходят через клеточные стенки соседних клеток, соединяя их между собой. Полость плазмодесм устлана плазмалеммой. Совокупность всех клеток, объединенных плазмодесмами, называется симпластом, между ними возможен регулируемый транспорт веществ.
Осмотическое давление в клетке
Осмотическое давление — избыточное гидростатическое давление на раствор, отделённый от чистого растворителя полупроницаемой мембраной, при котором прекращается диффузия растворителя через мембрану (осмос). Это давление стремится уравнять концентрации обоих растворов вследствие встречной диффузии молекул растворённого вещества и растворителя.
Мера градиента осмотического давления, то есть различия водного потенциала двух растворов, разделённых полупроницаемой мембраной, называется тоничностью. Раствор, имеющий более высокое осмотическое давление по сравнению с другим раствором, называется гипертоническим, имеющий более низкое — гипотоническим.
Тургор тканей — напряжённое состояние оболочек живых клеток. Тургорное давление — внутреннее давление, которое развивается в растительной клетке, когда в неё в результате осмоса входит вода и цитоплазма прижимается к клеточной стенке; это давление препятствует дальнейшему проникновению воды в клетку.
Тургор обуславливается тремя факторами: внутренним осмотическим давлением клетки, которое вызывает напряжение клеточной оболочки, внешним осмотическим давлением, а также упругостью клеточной оболочки.
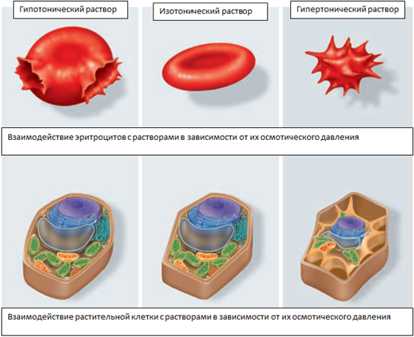
Рисунок 11. Взаимодействие эритроцитов и растительной клетки с растворами.
Дифференцировка клеток многоклеточного организма
Многоклеточные организмы состоят из клеток, которые в той или иной степени отличаются по строению и функциям, например, у взрослого человека около 230 различных типов клеток. Все они являются потомками одной клетки — зиготы (в случае полового размножения) — и приобретают различия в результате процесса дифференцировки.
Дифференцировка в подавляющем большинстве случаев не сопровождается изменением наследственной информации клетки, а обеспечивается лишь путем регуляции активности генов, специфический характер экспрессии генов наследуется во время деления материнской клетки обычно благодаря эпигенетическим механизмам. Однако есть исключения: например, при образовании клеток специфической иммунной системы позвоночных происходит перестройка некоторых генов, эритроциты млекопитающих полностью теряют всю наследственную информацию, а половые клетки — её половину.
Различия между клетками на первых этапах эмбрионального развития появляются, во-первых, вследствие неоднородности цитоплазмы оплодотворенной яйцеклетки, из-за чего во время процесса дробления образуются клетки, различающиеся по содержанию определенных белков и РНК; во-вторых, важную роль играет микроокружение клетки — её контакты с другими клетками и средой.
Возникновение клеток
Доподлинно неизвестно, когда на Земле появилась первая клетка и каким путем она возникла. Наиболее ранние вероятные ископаемые остатки клеток, приблизительный возраст которых оценен в 3,49 млрд лет, найдены на востоке Пилбары (Австралия), хотя биогенность их происхождения была поставлена под сомнение. О существовании жизни в раннем архее свидетельствуют также строматолиты того же периода.
Возникновению первых клеток должно было предшествовать накопление органических веществ в среде и появление определенной формы пребиотического метаболизма. Протоклетки содержали как минимум два обязательных элемента: наследственную информацию в виде молекул, способных к саморепликации, и определенного рода оболочки, которая ограждала внутреннее содержимое первых клеток от окружающей среды.
Наиболее вероятным кандидатом на роль саморепликативных молекул является РНК, поскольку она может одновременно выступать и носителем наследственной информации, и катализатором; кроме того, РНК, в отличие от ДНК, самодостаточна для осуществления биосинтеза белков.
Подробнее о клетке вы можете узнать из видео:
Без клетки нет жизни, клетка — это наша жизнь. Поэтому если узнавать больше о клетке, то можно объяснить, например, действие многих компонентов на нашу жизнь и самочувствие. Изучайте строение клетки и особенно важно изучать клетку будущим врачам.
6.Происхождение, строение и функции клеточной оболочки.
История открытия клетки, клеточная теория. Клетка - основная структурная единица живого. Открытие ее (Гук,1665; Мальпиги; Грю, 1671) связано с изобретением светового микроскопа. Дальнейшие исследования Р. Вирхова, К. Бэра показали, что организмы начинают свое развитие из одной клетки, каждая клетка образуется путем деления материнской. Это нашло свое выражение в клеточной теории, основные положения которой сформулированы французским ботаником Дютроше (1824), русским ботаником Горяниновым П.Ф.(1834) и немецкими исследователями Шлейденом и Шванном (1838-1839).
Современная клеточная теория включает следующие основные положения:
1 .Клетка - основная единица строения, развития и жизнедеятельности живых организмов.
2. Клетки растений и животных сходны по строению.
3. Клетки образуются в результате деления материнских клеток.
4. Клетки специализированы по функциям и образуют ткани.
5. Ткани формируют органы.
Более глубокие представления о строении клетки связаны с появлением фазово-контрастного, электронного, трансмиссионного и сканирующего микроскопов, которые обеспечивают увеличение в сотни тысяч раз.
Строение клетки. Основными структурными частями клетки являются: оболочка, цитоплазма, ядро, вакуоль. Живая часть клетки (цитоплазма, ядро) называется протопластом.
Клеточная оболочка. Клетки растений окружены плотной оболочкой. Наличием ее они отличаются от клеток животных, хотя установлено, что и клетки животных покрыты оболочкой гликопротеидного вещества муцина (сахар и белок) - яйца морских ежей, амфибий, клетки, выстилающие желудочно-кишечный тракт, эпителий и др.
Клеточная оболочка защищает протопласт от неблагоприятных внешних воздействий и придает клетке определенную форму и прочность.
Клеточная оболочка состоит главным образом из полисахаридов - целлюлозы 50%, гемицеллюлозы 30% и пектиновых веществ 20%.
Целлюлоза имеет фибриллярное строение. Глюкозные остатки в молекуле целлюлозы образуют цепи - мицеллы, которые объединяются в пучки. Мелкие пучки в крупные и т.д.
Чистая целлюлоза бесцветна, прочна и стойка против различного рода механических и физических воздействий. Промежутки между пучками мицелл заполнены пектиновыми веществами, способными при намачивании набухать. Пектиновые вещества заполняют и межклеточные пространства, склеивая клетки между собой. Часто на стенках клеток откладывается не целлюлоза, а гемицеллюлоза, вещество, стоящее ближе к крахмалу.
Утолщение клеточных оболочек происходит в основном за счет пропитывания их особыми веществами, обеспечивающими дополнительную прочность и стойкость. Это лигнин, суберин, кутин. Лигнин - вещество, близкое к целлюлозе, но углерода в нем относительно больше. Такое видоизменение - одревеснение.
Суберин и кутин по своей природе близки к жирам. Клеточные оболочки, пропитанные ими, не смачиваются водой и почти непроницаемы для воды и газов. Это уменьшает испарение с поверхности клеток. Кутикулой покрывается только наружная поверхность клеточных оболочек (поверхность листа), поэтому клетки сохраняют свою жизнеспособность. Сквозное пропитывание этими веществами клеточной оболочки приводит к опробковению, что вызывает отмирание протопласта клетки.
Минерализация оболочек отмечена в небольшой степени у всех клеток. Обычно это бывают соли кальция или кремниевой кислоты. Кальций встречается в виде углекислой или щавелевокислой извести. Углекислая известь может откладываться не только в оболочках, но на поверхности эпидермы, осаждаясь из выделений водных устьиц (в жгучих волосках крапивы вместе с кремнеземом).
Формирование и рост клеточной оболочки. Образовавшаяся в процессе деления клеток общая для них перегородка представляет собой тончайшую пленку из целлюлозы. В нее даже проникают частицы цитоплазмы протопластов соседних клеток. Эта тонкая целлюлозная пленка называется первичной оболочкой, содержит около 5% целлюлозы. Вначале первичная оболочка общая для двух соседних клеток. Затем она утолщается и разделяется, и каждая клетка получает свою первичную оболочку. Между ними возникает тончайший слой аморфного вещества - межклеточная или срединная пластинка. По физическим и химическим свойствам она имеет пектиновую природу. Она может разрушаться, и соседние клетки разъединяются. По мере роста молодых клеток увеличивается и первичная оболочка, становится толще.
Вначале, у очень молодых клеток мицеллярные тяжи образуют трехмерную сетку. Она легко растягивается. По мере роста клетки, оболочка растягивается, к уже имеющимся пучкам мицелл присоединяются новые. Сетка становится более плотной и тесной. Общая толщина ее растет. Пластичность оболочки снижается, и закрепляется определенный размер и форма клетки. Последующее утолщение клеточной оболочки называется вторичным, а наслаивающаяся оболочка называется вторичной оболочкой. Такова схематично сложная морфологическая структура клеточной оболочки. Сложность эта усугубляется еще тем, что вторичное утолщение никогда не бывает сплошным, равномерным, а бывает самым разнообразным: кольчатым, спиральным, лесничным, сетчатым, точечным.
Кольчатые и спиральные утолщения представляют собой кольца или спирали круглого сечения, расположенные внутри клетки, имеющей форму цилиндрической трубки. Они соединены с внутренней поверхностью первичной оболочки лишь узкой спайкой, не препятствуют удлинению клеток, росту их в длину.
Утолщения могут быть в виде сетки на внутренней поверхности клеточной оболочки, и в виде ступенек, вдающихся внутрь клетки, и почти сплошное. В последнем случае остаются не утолщенными лишь узкие пространства округлой или щелевидной формы.
В первичной оболочке имеются не утолщенные места - поры. В связи с различным характером утолщений, форма и конструкция пор может быть самой разнообразной:
Простые поры - у них стенки канала, образуемого вторичной оболочкой, опускаются к первичной оболочке ровно, отвесно.
Полуокаймленные - вторичная оболочка с одной стороны.
Окаймленные поры - вторичная оболочка как бы нависает над не утолщенным местом так, что канал во вторичной оболочке приобретает форму воронки, приставленной раструбом к первичной оболочке. Пленка первичной оболочки, разгораживающая в поре два встречных канала, может иметь утолщение в виде диска или линзы, которое называется торусом.
Очертания как простой, так и окаймленной поры не всегда округлое, оно может быть вытянутым, эллиптическим. Первичная оболочка в области пор пронизана тончайшими отверстиями-перфорациями, через которые проходят тяжи цитоплазмы-плазмодесмы, соединяющие клетки и обеспечивающие жизнедеятельность растения как целостного организма.
Видоизменения клеточной оболочки:
одревеснение – инкрустация лигнином;
опробковение – инкрустация суберином;
кутинизация – образование слоя кутина на внешней поверхности оболочки;
минерализация – пропитывание солями кальция или кремния.
Ослизнение клеточных оболочек. Может быть нормальное (биологическое), полезное для растений и патологическое (болезненное), вызываемое бактериями.
Нормальное ослизнение целлюлозных оболочек поверхностного слоя клеток семян льна, айвы, тыквы, некоторых видов ромашки способствует закреплению семян в почве, создается лучший контакт с ней и лучшие условия произрастания.
В патологических случаях могут ослизняться стенки не только поверхностных, но и глубинных клеток. Этот процесс вызывается специальными бактериями, является болезнью растений и называется гуммозом. Часто поражает плодовые деревья, особенно вишни, сливы. Из коры дерева вытекает слизь, вишневый клей. Гуммоз развивается медленно и в конечном счете приводит к гибели дерева.
Клетка — Википедия
Кле́тка — структурно-функциональная элементарная единица строения и жизнедеятельности всех организмов. Обладает собственным обменом веществ, способна к самовоспроизведению. Организм, состоящий из одной клетки, называется одноклеточным (многие простейшие и бактерии). Раздел биологии, занимающийся изучением строения и жизнедеятельности клеток, называется цитологией. Также принято говорить о биологии клетки, или клеточной биологии.
Срез пробкового дерева из книги Роберта Гука «Микрография», 1635—1703 Первое наблюдение за клетками, с использованием раннего микроскопа[1]. Это привело к развитию теории клеток.Первым человеком, увидевшим клетки, был английский учёный Роберт Гук (известный открытием закона Гука). В 1665 году, пытаясь понять, почему пробковое дерево хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему соты в ульях медоносных пчёл, и он назвал эти ячейки клетками (по-английски cell означает «ячейка, клетка»).
В 1675 году итальянский врач Марчелло Мальпиги подтвердил клеточное строение растений, а в 1681 году — английский ботаник Неемия Грю. О клетке стали говорить как о «пузырьке, наполненном питательным соком». В 1674 году голландский мастер Антоний ван Левенгук с помощью микроскопа впервые увидел в капле воды «зверьков» — движущиеся живые организмы (инфузории, амёбы, бактерии). Также Левенгук впервые наблюдал животные клетки — эритроциты и сперматозоиды. Таким образом, к началу XVIII века учёные знали, что под большим увеличением растения имеют ячеистое строение, и видели некоторые организмы, которые позже получили название одноклеточных. В 1802—1808 годах французский исследователь Шарль-Франсуа Мирбель установил, что растения состоят из тканей, образованных клетками. Ж. Б. Ламарк в 1809 году распространил идею Мирбеля о клеточном строении и на животные организмы. В 1825 году чешский учёный Я. Пуркине открыл ядро яйцеклетки птиц, а в 1839 ввёл термин «протоплазма». В 1831 году английский ботаник Р. Броун впервые описал ядро растительной клетки, а в 1833 году установил, что ядро является обязательным органоидом клетки растения. С тех пор главным в организации клеток считается не оболочка, а содержимое.
Клеточная теория[править | править код]
Клеточная теория строения организмов была сформирована в 1839 году немецкими учёными, зоологом Теодором Шванном и ботаником М. Шлейденом, и включала в себя три положения. В 1858 году Рудольф Вирхов дополнил её ещё одним положением, однако в его идеях присутствовал ряд ошибок: так, он предполагал, что клетки слабо связаны друг с другом и существуют каждая «сама по себе». Лишь позднее удалось доказать целостность клеточной системы.
В 1878 году русским учёным И. Д. Чистяковым открыт митоз в растительных клетках; в 1878 году В. Флемминг и П. И. Перемежко обнаруживают митоз у животных. В 1882 году В. Флемминг наблюдает мейоз у животных клеток, а в 1888 году Э. Страсбургер — у растительных.
Клеточная теория является одной из основополагающих идей современной биологии, она стала неопровержимым доказательством единства всего живого и фундаментом для развития таких дисциплин, как эмбриология, гистология и физиология. На сегодняшний день теория содержит такие утверждения:
- Клетка — элементарная единица строения, функционирования, размножения и развития всех живых организмов. Вне клетки нет жизни.
- Клетка — целостная система, содержащая большое количество связанных друг с другом элементов — органелл.
- Клетки различных организмов похожи (гомологичны) по строению и основным свойствам и имеют общее происхождение.
- Увеличение количества клеток происходит путём их деления, после репликации их ДНК: клетка — от клетки.
- Многоклеточный организм — система из большого количества клеток, объединённых в системы тканей и органов, связанных между собой гуморальной и нервной регуляциями.
- Клетки многоклеточных организмов обладают одинаковым полным фондом генетического материала этого организма, всеми возможными потенциями для проявления этого материала, — но отличаются по уровню экспрессии (работы) отдельных генов, что приводит к их морфологическому и функциональному разнообразию — дифференцировке[2].
Количество и формулировки отдельных положений современной клеточной теории в разных источниках могут отличаться.
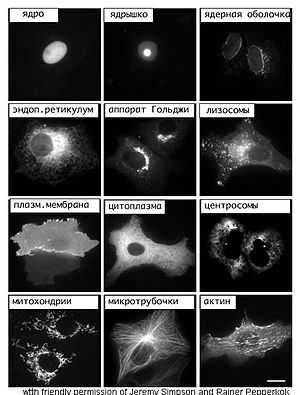
Впервые клетки удалось увидеть только после создания оптических (световых) микроскопов. С того времени микроскопия остается одним из важнейших методов исследования клеток. Световая микроскопия, несмотря на небольшое разрешение, позволяла наблюдать за живыми клетками. В XX веке была изобретена электронная микроскопия, которая позволила изучить ультраструктуру клеток.
Для изучения функций клеток и их частей используют разнообразные биохимические методы — как препаративные, например фракционирование методом дифференциального центрифугирования, так и аналитические. Для экспериментальных и практических целей используют методы . Все упомянутые методические подходы могут использоваться в сочетании с методами культуры клеток.
Оптическая микроскопия[править | править код]
В оптическом микроскопе увеличение объекта достигается благодаря серии линз, через которые проходит свет. Максимальное увеличение составляет более 1000 раз. Также важной характеристикой является разрешение — расстояние между двумя точками, которые ещё распознаются отдельно. Разрешение характеризует чёткость изображения. Эта величина ограничивается длиной световой волны, и даже при использовании самого коротковолнового света — ультрафиолетового — можно достичь разрешения только около 200 нм; такое разрешение было получено ещё в конце XIX века. Малейшие структуры, которые можно наблюдать под оптическим микроскопом, это митохондрии и бактерии. Их линейный размер составляет примерно 500 нм. Однако объекты размером меньше 200 нм видны в световом микроскопе, если они сами излучают свет. Эта особенность используется в флуоресцентной микроскопии, когда клеточные структуры или отдельные белки связываются со специальными флуоресцентными белками или антителами с флуоресцентными метками. На качество изображения, полученного с помощью оптического микроскопа, влияет также контрастность — её можно увеличить, используя различные методы окраски клеток. Для изучения живых клеток используют фазово-контрастную, дифференциальную интерференционно-контрастную и темнопольную микроскопию. Конфокальные микроскопы позволяют улучшить качество флуоресцентных изображений[3][4].
Изображения, полученные с помощью оптической микроскопииЭлектронная микроскопия[править | править код]
В 30-х годах XX века был сконструирован электронный микроскоп, в котором вместо света через объект пропускается пучок электронов. Теоретический предел разрешения для современных электронных микроскопов составляет около 0,002 нм, однако из практических причин для биологических объектов достигается разрешение только около 2 нм. С помощью электронного микроскопа можно изучать ультраструктуру клеток. Различают два основных типа электронной микроскопии: сканирующую и трансмиссионную. Сканирующая (растровая) электронная микроскопия (РЭМ) используется для изучения поверхности объекта. Образцы зачастую покрывают тонкой плёнкой золота. РЭМ позволяет получать объёмные изображения. Трансмиссионная (просвечивающая) электронная микроскопия (ПЭМ) — используется для изучения внутреннего строения клетки. Пучок электронов пропускается через объект, предварительно обработанный тяжёлыми металлами, которые накапливаются в определённых структурах, увеличивая их электронную плотность. Электроны рассеиваются на участках клетки с большей электронной плотностью, в результате чего на изображениях эти области выглядят темнее[3][4].
Фракционирование клеток[править | править код]
Для установления функций отдельных компонентов клетки важно выделить их в чистом виде, чаще всего это делается с помощью метода дифференциального центрифугирования. Разработаны методики, позволяющие получить чистые фракции любых клеточных органелл. Получение фракций начинается с разрушения плазмалеммы и образования клеток. Гомогенат последовательно центрифугируется при различных скоростях, на первом этапе можно получить четыре фракции: (1) ядер и крупных обломков клеток, (2) митохондрий, пластид, лизосом и пероксисом, (3) — пузырьков аппарата Гольджи и эндоплазматического ретикулума, (4) рибосом, в останутся белки и более мелкие молекулы. Дальнейшее дифференциальное центрифугирование каждой из смешанных фракций позволяет получить чистые препараты органелл, к которым можно применять разнообразные биохимические и микроскопические методы[2].
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
- прокариоты (доядерные) — более простые по строению, возникли в процессе эволюции раньше;
- эукариоты (ядерные) — более сложные, возникли позже. Клетки, составляющие тело человека, являются эукариотическими.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам.
Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органеллы и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждая из органелл клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
Прокариотическая клетка[править | править код]
Прокариоты (от др.-греч. πρό — ‘перед’, ‘до’ и κάρῠον — ‘ядро’) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
Эукариотическая клетка[править | править код]
Эукариоты (эвкариоты, от др.-греч. εὖ ‘хорошо’, ‘полностью’ и κάρῠον — ‘ядро’) — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот есть система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.
Фимбрии кишечной палочки, которые позволяют ей прикрепляться к субстрату (ОМ)Клетки двух основных групп прокариот — бактерий и архей — похожи по структуре, характерными их признаками являются отсутствие ядра и мембранных органелл.
Основными компонентами прокариотической клетки являются:
- Клеточная стенка, которая окружает клетку извне, защищает её, придаёт устойчивую форму, предотвращающую от осмотического разрушения. У бактерий клеточная стенка состоит из пептидогликана (муреина), построенного из длинных полисахаридных цепей, соединённых между собой короткими пептидными перемычками. По строению клеточной стенки различают две группы бактерий:
- Клеточная стенка архей не содержит муреина, а построена в основном из разнообразных белков и полисахаридов[4].
- Капсула — имеющаяся у некоторых бактерий слизистая оболочка, расположенная снаружи от клеточной стенки. Состоит в основном из разнообразных белков, углеводов и уроновых кислот. Капсулы защищают клетки от высыхания, могут помогать бактериям в колониях удерживаться вместе, а индивидуальным бактериям — прикрепляться к различным субстратам. Кроме этого, капсулы предоставляют клетке дополнительную защиту: например, капсулированные штаммы пневмококков свободно размножаются в организме и вызывают воспаление лёгких, тогда как некапсулированные быстро уничтожаются иммунной системой и являются абсолютно безвредными[5].
- Пили или ворсинки — тонкие волоскоподобные выросты, что присутствуют на поверхности бактериальных клеток. Существуют различные типы пилей, из которых наиболее распространёнными являются:
- Жгутики — органеллы движения некоторых бактерий. Бактериальный жгутик построен значительно проще эукариотического, и он в 10 раз тоньше, не покрыт плазматической мембраной и состоит из одинаковых молекул белков, которые образуют цилиндр. В мембране жгутик закреплён при помощи базального тела[4].
- Плазматическая и внутренние мембраны. Общий принцип устройства клеточных мембран не отличается от эукариот, однако в химическом составе мембраны есть немало различий, в частности, в мембранах прокариот отсутствуют молекулы холестерина и некоторых липидов, присущих мембранам эукариот. Большинство прокариотических клеток (в отличие от эукариотических) не имеют внутренних мембран, которые разделяют цитоплазму на отдельные компартменты. Только у некоторых фотосинтетических и аэробных бактерий плазмалемма образует вгибание внутрь клетки, что выполняет соответствующие метаболические функции[4].
- Нуклеоид — не ограниченный мембранами участок цитоплазмы, в котором расположена кольцевая молекула ДНК — «бактериальная хромосома», где хранится весь генетический материал клетки[4].
- Плазмиды — небольшие дополнительные кольцевые молекулы ДНК, несущие обычно всего несколько генов. Плазмиды, в отличие от бактериальной хромосомы, не являются обязательным компонентом клетки. Обычно они придают бактерии определённые полезные для неё свойства, такие как устойчивость к антибиотикам, способность усваивать из среды определённые энергетические субстраты, способность инициировать половой процесс и т. д.[4][5]
- Рибосомы прокариот, как и у всех других живых организмов, отвечают за осуществление процесса трансляции (одного из этапов биосинтеза белка). Однако бактериальные рибосомы несколько меньше, чем эукариотические (коэффициенты седиментации 70S и 80S соответственно), и имеют другой состав белков и РНК. Из-за этого бактерии, в отличие от эукариот, чувствительны к таким антибиотикам, как эритромицин и тетрациклин, которые избирательно действуют на 70S-рибосомы[5].
- Эндоспоры — окружённые плотной оболочкой структуры, содержащие ДНК бактерии и обеспечивающее выживание в неблагоприятных условиях. К образованию эндоспор способны лишь некоторые виды прокариот, например представители родов Clostridium (C. tetani — возбудитель столбняка, C. botulinum — возбудитель ботулизма, C. perfringens — возбудитель газовой гангрены и т. п.) и Bacillus (в частности B. anthracis — возбудитель сибирской язвы). Для образования эндоспоры клетка реплицирует свою ДНК и окружает копию плотной оболочкой, из созданной структуры удаляется избыток воды, и в ней замедляется метаболизм[4]. Споры бактерий могут выдерживать довольно жёсткие условия среды, такие как длительное высушивание, кипячение, коротковолновое облучение и др.[5]
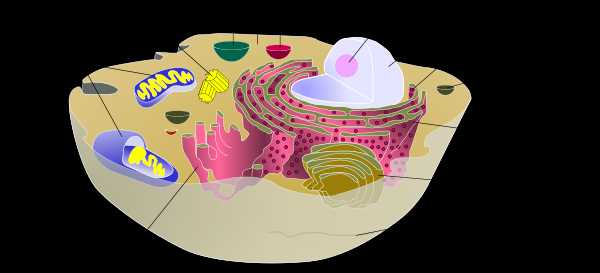 Схематическое изображение животной клетки (подписи составных частей — ссылки на статьи про них).
Схематическое изображение животной клетки (подписи составных частей — ссылки на статьи про них). Поверхностный комплекс животной клетки[править | править код]
Состоит из гликокаликса, плазмалеммы и расположенного под ней кортикального слоя цитоплазмы. Плазматическая мембрана называется также плазмалеммой, наружной клеточной мембраной, цитолеммой и т. д. Её толщина — около 10 нанометров. Обеспечивает разграничение клетки и внешней среды, а также пропускание внутрь и наружу некоторых веществ.
На сохранение целостности своей мембраны клетка не тратит энергии: молекулы удерживаются по тому же принципу, по которому удерживаются вместе молекулы жира — гидрофобным частям молекул термодинамически выгоднее располагаться в непосредственной близости друг к другу. Гликокаликс представляет собой «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов, гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции.
Плазматическая мембрана животных клеток в основном состоит из фосфолипидов и липопротеидов со вкрапленными в неё молекулами белков, в частности, поверхностных антигенов и рецепторов.
В кортикальном (прилегающем к плазматической мембране) слое цитоплазмы находятся специфические элементы цитоскелета — упорядоченные определённым образом актиновые микрофиламенты. Основной и самой важной функцией кортикального слоя (кортекса) являются псевдоподиальные реакции: выбрасывание, прикрепление и сокращение псевдоподий. При этом микрофиламенты перестраиваются, удлиняются или укорачиваются. От структуры цитоскелета кортикального слоя зависит также форма клетки (например, наличие микроворсинок).
Структура цитоплазмы[править | править код]
Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органеллы. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами», и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
Рибосомы[править | править код]
Рибосомы — органоиды, необходимые клетке для синтеза белка. Их размер составляет примерно 20—30 нм. В клетке их насчитывается несколько миллионов. Рибосомы образованы из двух субъединиц: большой и малой
Клетка
На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.
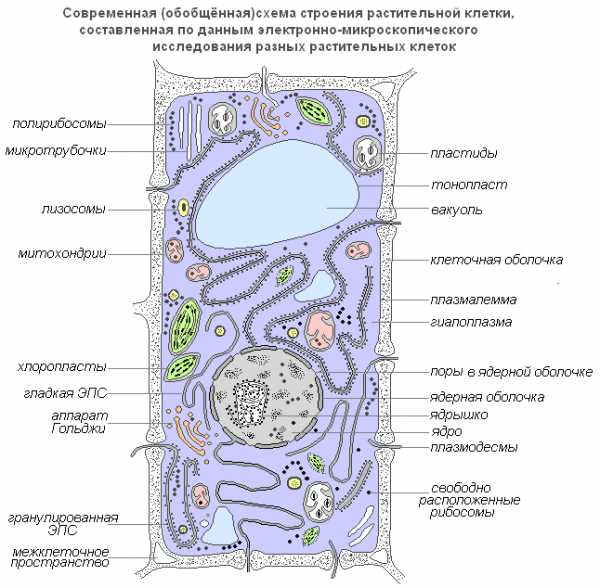
Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.
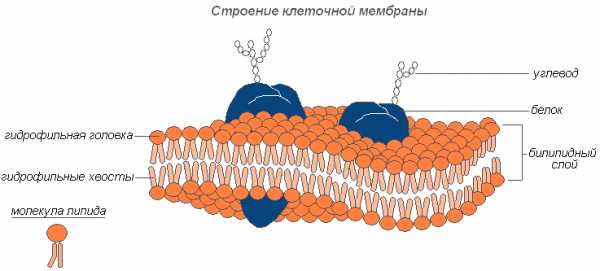
Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования – органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.
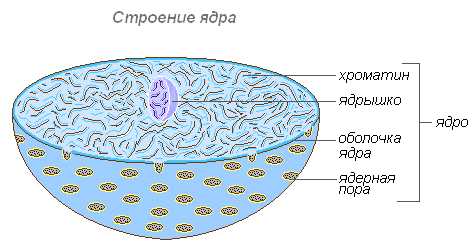
Строение ядра
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.
Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).
Строение лизосомы
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.
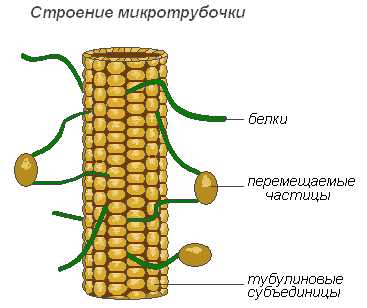
Строение микротрубочки
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.
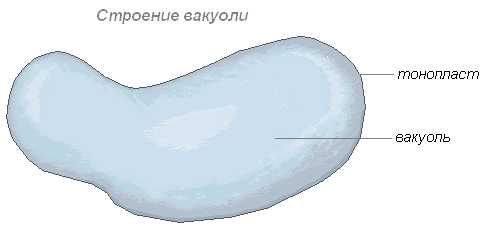
Строение вакуоли
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.
Строение хлоропласта
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.
Строение лейкопласта
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.
Строение хромопласта
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.
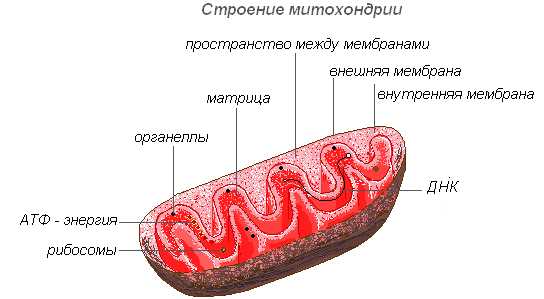
Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.
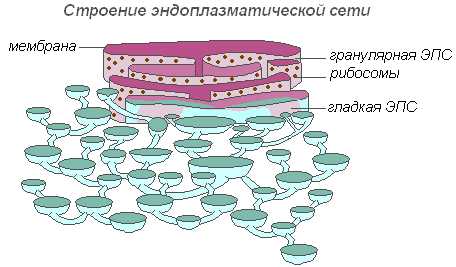
Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.
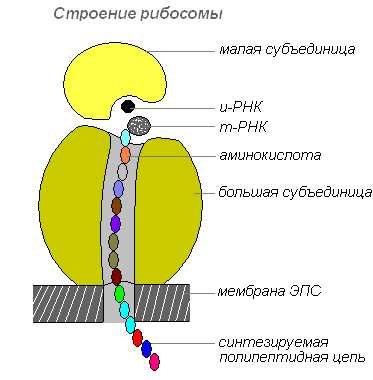
Строение рибосомы
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
Клеточная стенка — Википедия
Материал из Википедии — свободной энциклопедии
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 19 декабря 2018; проверки требуют 3 правки. Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 19 декабря 2018; проверки требуют 3 правки. Растительная клетка. Зелёная оболочка — клеточная стенка.Клеточная стенка — оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, архей, грибов и растений. Животные и многие простейшие не имеют клеточной стенки.
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного. Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми и липотейхоевыми кислотами. При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов состоят из хитина и глюканов.
Большинство водорослей имеют клеточную стенку из целлюлозы и различных гликопротеинов. Включения дополнительных полисахаридов имеют большое таксономическое значение.
Диатомовые водоросли синтезируют свою клеточную стенку из кремнезёма.
Важнейшей отличительной особенностью растительной клетки является наличие прочной клеточной стенки, основным компонентом которой является целлюлоза. Клеточная стенка высших растений представляет собой сложноорганизованный, преимущественно полимерный, внеклеточный матрикс, окружающий каждую клетку. Растительная клетка, лишённая клеточной стенки, обозначается термином протопласт. В клеточных стенках растений существуют углубления — поры, через которые проходят цитоплазматические канальца — плазмодесмы, осуществляющие контакт соседних клеток и обмен веществами между ними.
Химический состав и пространственная организация полимеров клеточной стенки отличаются у разных видов, клеток разных тканей одного растения и иногда у разных частей стенки вокруг одного протопласта.
Кроме того, строение клеточной стенки изменяется в онтогенезе растительного организма. Первичная клеточная стенка формируется при делении и сохраняется во время роста клетки. Формирование вторичной клеточной стенки происходит с внутренней стороны от первичной стенки и связано с окончанием роста и специализацией (дифференцировкой) клеток растения. Снаружи от первичной клеточной стенки, между первичными стенками двух соседних клеток, располагается срединная пластинка (состоит преимущественно из кальциевых и магниевых солей пектиновых веществ).
Строение первичной клеточной стенки растений.Первичная клеточная стенка высших растений состоит из трёх взаимодействующих, но структурно независимых трехмерных сетей полимеров. Основная сеть состоит из фибрилл целлюлозы и связывающих их гемицеллюлоз (или сшивочных гликанов). Вторая сеть состоит из пектиновых веществ. Третья сеть представлена, как правило, структурными белками клеточной стенки. Также следует отметить, что у растений клады commelinids (группа в системах APG) и у представителей семейства Маревые первичная клеточная стенка содержит значительное количество ароматических веществ (гидроксикоричные кислоты, главным образом феруловая и п-кумаровая). При этом у представителей клады commelinids гидроксикоричные кислоты присоединяются к сшивочным гликанам (к глюкуроноарабиноксиланам), а у семейства Маревые к пектиновым веществам (к рамногалактуронанам I).
Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жёсткость клетки для структурной и механической поддержки, придают форму клетке, направление её роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургору, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают от патогенов, проникающих из окружающей среды, и запасают углеводы для растения.
| Органоиды клетки | Строение органоидов |
Функция |
Присутствие органоидов в клетках | |
растений |
животных |
|||
| Хлоропласт | Представляет собой разновидность пластид. | Окрашивает растения в зеленый цвет, в нем происходит фотосинтез | + |
– |
| Лейкопласт | оболочка состоит из двух элементарных мембран, внутренняя из них, врастая в строму, образует немногочисленные тилакоиды. | Окрашивает растения в желтый цвет, синтезирует и накапливает крахмал. | + |
– |
| Хромопласт | пластид с жёлтой, оранжевой и красной окраской, окраска обусловлена пигментами – каротиноидами | Бесцветное окрашивание растения | + |
– |
| Вакуоль | Занимает до 90 % объема зрелой клетки, заполнена клеточным соком | Функция питания | + |
– |
| Микротрубочки | Состоят из белка тубулина, расположены около плазматической мембраны | Участвуют в отложении целлюлозы на клеточных стенках, участвуют в перемещении в цитоплазме различных органоидов. При делении клетки микротрубочки составляют основу структуры веретена деления | + |
+ |
| Плазматическая мембрана | Состоит из билипидного слоя, пронизанного белками, погруженными на различную глубину. | Барьер, транспорт веществ, сообщение клеток между собой | + |
+ |
| Гладкий ЭПР | Система плоских и ветвящихся трубочек. | Осуществляет синтез и выделение липидов | + |
+ |
| Шероховатый ЭПР | Название получил из–за множества рибосом, находящихся на его поверхности | Синтез белков, их накопление и преобразование для выделения из клетки наружу | + |
+ |
| Ядро | Окружено двойной ядерной мембраной, имеющей поры. Наружная ядерная мембрана образует непрерывную структуру с мембраной ЭПР. Содержит одно или несколько ядрышек. | Носитель наследственной информации, центр регуляции активности клетки. | + |
+ |
| Клеточная стенка | Состоит из длинных молекул целлюлозы, собранных в пучки, называемые микрофибриллами. | Внешний каркас, или защитную оболочку, обеспечивает тургор растительных клеток | + |
+ |
| Плазмодесмы | Мельчайшие цитоплазматические каналы, которые пронизывают клеточные стенки. | Объединяют протопласты соседних клеток | + |
– |
| Митохондрии | Содержат ферменты для синтеза АТФ. Внутренняя мембрана митохондрий образует многочисленные складки. | Аккумулятор энергии, осуществляет аэробное дыхание. | + |
+ |
| Аппарат Гольджи | Состоит из стопки плоских мешочков, называемых цистернами | Синтезирует полисахариды | + |
+ |
| Лизосомы | Пузырьки, содержащие концентрированные гидролитические ферменты, которые становятся активными в кислой среде | Участвуют в растворении веществ, попавших в клетку | + |
+ |
| Рибосомы | Состоит из двух неравных субъединиц – большой и малой, на которые может диссоциировать. | Место биосинтеза белка | + |
+ |
| Эндоцитозный пузырек | Содержит слишком большие молекулы | Содержит слишком большие молекулы, которые не могут проникнуть через мембрану способами диффузии или активного транспорта | – |
+ |
| Цитоплазма | Состоит из воды с большим количеством растворенных в ней веществ, содержащих глюкозу, белки и ионы. | В ней расположены другие органоиды клетки | + |
+ |
| Микрофиламенты | Волокна из белка актина, обычно располагаются пучками вблизи от поверхности клеток. | Играют важную роль в подвижности клеток | – |
+ |
| Секреторный пузырек | много в клетках, активно синтезирующих вещества, например, в клетках островков Лангерганса | Выносит вещества за пределы клетки | – |
+ |
| Центриоли | Могут входить в состав митотического аппарата клетки. В диплоидной клетке содержится две пары центриолей. | Участвуют в процессе деления клетки у животных | – |
+ |
| Пероксисома | Группа пузырьков, известных как микротела | Важна для замедления старения клеток | – |
+ |
| Микроворсинки | Выступы плазматической мембраны | Увеличивают наружную поверхность клетки, микроворсинки в совокупности образуют кайму клетки | – |
+ |
11.Клеточная стенка растений. Строение и функции – оболочки клеток растений, животных и прокариот, сравнение.
По форме клетки необычайно разнообразны: одни имеют округлую форму, другие похожи на звездочки со многими лучами, третьи вытянутые и т.д. Различны клетки и по размеру – от мельчайших, с трудом различимых в световом микроскопе, до прекрасно видимых невооруженным глазом (например, икринки рыб и лягушек).
Прокариоты(от лат.pro– перед, раньше, вместо и греч.karyon– ядро) – это организмы, клетки которых не имеют ограниченного мембраной ядра, т.е. все бактерии, включая архебактерии и цианобактерии. Общее число видов прокариот около 6000. Вся генетическая информация прокариотической клетки (генофор) содержится в одной-единственной кольцевой молекуле ДНК. Митохондрии и хлоропласты отсутствуют, а функции дыхания или фотосинтеза, обеспечивающие клетку энергией, выполняет плазматическая мембрана. Размножаются прокариоты без выраженного полового процесса путем деления надвое. Прокариоты способны осуществлять целый ряд специфических физиологических процессов: фиксируют молекулярный азот, осуществляют молочнокислое брожение, разлагают древесину, окисляют серу и железо.
Эукариоты– это высшие организмы, имеющие четко оформленное ядро, которое оболочкой отделяется от цитоплазмы (кариомембраной). К эукариотам относятся все высшие животные и растения, а также одноклеточные и многоклеточные водоросли, грибы и простейшие. Ядерная ДНК у эукариот заключена в хромосомах. Эукариоты обладают клеточными органоидами, ограниченными мембранами.
Отличия эукариот от прокариот
– Эукариоты имеют настоящее ядро: генетический аппарат эукариотической клетки защищен оболочкой, схожей с оболочкой самой клетки. – Включенные в цитоплазму органоиды окружены мембраной.
Строение клеток растений и животных
Клетка любого организма представляет собой сис-тему. Она состоит из трех взаимосвязанных между собой частей: оболочки, ядра и цитоплазмы.
| Органоиды клетки | Строение органоидов | Функция | Присутствие органоидов в клетках | |
| растений | животных | |||
| Хлоропласт | Представляет собой разновидность пластид | Окрашивает растения в зеленый цвет, в нем происходит фотосинтез | + | – |
| Лейкопласт | Оболочка состоит из двух элементарных мембран; внутренняя, врастая в строму, образует немногочисленные тилакоиды | Синтезирует и накапливает крахмал, масла, белки | + | – |
| Хромопласт | Пластиды с желтой, оранжевой и красной окраской, окраска обусловлена пигментами – каротиноидами | Красная, желтая окраска осенних листьев, сочных плодов и др. | + | – |
| Вакуоль | Занимает до 90% объема зрелой клетки, заполнена клеточным соком | Поддержание тургора, накопление запасных веществ и продуктов обмена, регуляция осмотического давления и др. | + | – |
| Микротрубочки | Состоят из белка тубулина, расположены около плазматической мембраны | Участвуют в отложении целлюлозы на клеточных стенках, перемещении в цитоплазме различных органоидов. При делении клетки микротрубочки составляют основу структуры веретена деления | + | + |
| Плазматическая мембрана (ЦПМ) | Состоит из липидного бислоя, пронизанного белками, погруженными на различную глубину | Барьер, транспорт веществ, сообщение клеток между собой | + | + |
| Гладкий ЭПР | Система плоских и ветвящихся трубочек | Осуществляет синтез и выделение липидов | + | + |
| Шероховатый ЭПР | Название получил из-за множества рибосом, находящихся на его поверхности | Синтез белков, их накопление и преобразование для выделения из клетки наружу | + | + |
| Ядро | Окружено двойной ядерной мембраной, имеющей поры. Наружная ядерная мембрана образует непрерывную структуру с мембраной ЭПР. Содержит одно или несколько ядрышек | Носитель наследственной информации, центр регуляции активности клетки | + | + |
| Клеточная стенка | Состоит из длинных молекул целлюлозы, собранных в пучки, называемые микрофибриллами | Внешний каркас, защитная оболочка | + | – |
| Плазмодесмы | Мельчайшие цитоплазматические каналы, которые пронизывают клеточные стенки | Объединяют протопласты соседних клеток | + | – |
| Митохондрии | Содержат ферменты для синтеза АТФ. Внутренняя мембрана митохондрий образует многочисленные складки | Синтез АТФ (аккумуляция энергии) | + | + |
| Аппарат Гольджи | Состоит из стопки плоских мешочков – цистерн, или диктиосом | Синтез полисахаридов, формирование ЦПМ и лизосом | + | + |
| Лизосомы | Пузырьки, содержащие концентрированные гидролитические ферменты, которые становятся активными в кислой среде | Внутриклеточное пищеварение | – | + |
| Рибосомы | Состоят из двух неравных субъединиц – большой и малой, на которые могут диссоциировать | Место биосинтеза белка | + | + |
| Цитоплазма | Состоит из воды с большим количеством растворенных в ней веществ, содержащих глюкозу, белки и ионы | В ней расположены другие органоиды клетки и осуществляются все процессы клеточного метаболизма | + | + |
| Микрофиламенты | Волокна из белка актина, обычно располагаются пучками вблизи поверхности клеток | Участвуют в подвижности и изменении формы клеток | + | + |
| Центриоли | Могут входить в состав митотического аппарата клетки. В диплоидной клетке содержится две пары центриолей | Участвуют в процессе деления клетки у животных; в зооспорах водорослей, мхов и у простейших образуют базальные тельца ресничек | + - | + |
| Микроворсинки | Выступы плазматической мембраны | Увеличивают наружную поверхность клетки, микроворсинки в совокупности образуют кайму клетки | - | + |
Клеточная стенка, пластиды и центральная вакуоль присущи только растительным клеткам. 2. Лизосомы, центриоли, микроворсинки присутствуют в основном только в клетках животных организмов. 3. Все остальные органоиды характерны как для растительных, так и для животных клеток.
Строение оболочки клеток
Клеточная оболочка располагается снаружи клетки, отграничивая последнюю от внешней или внутренней среды организма. Ее основу составляет плазмалемма (клеточная мембрана) и углеводно-белковая составляющая.
Функции клеточной оболочки:
– поддерживает форму клетки и придает механическую прочность клетке и организму в целом; – защищает клетку от механических повреждений и попадания в нее вредных соединений; – осуществляет узнавание молекулярных сигналов; – регулирует обмен веществ между клеткой и средой; – осуществляет межклеточное взаимодействие в многоклеточном организме.
Функция клеточной стенки:
– представляет собой внешний каркас – защитную оболочку; – обеспечивает транспорт веществ (через клеточную стенку проходит вода, соли, молекулы многих органических веществ).
Наружный слой клеток животных, в отличие от клеточных стенок растений, очень тонкий, эластичный. Он не виден в световой микроскоп и состоит из разнообразных полисахаридов и белков. Поверхностный слой животных клеток называется гликокаликсом, выполняет функцию непосредственной связи клеток животных с внешней средой, со всеми окружающими ее веществами, опорной роли не выполняет.
Под гликокаликсом животной и клеточной стенкой растительной клетки расположена плазматическая мембрана, граничащая непосредственно с цитоплазмой. В состав плазматической мембраны входят белки и липиды. Они расположены упорядоченно за счет различных химических взаимодействий друг с другом. Молекулы липидов в плазматической мембране расположены в два ряда и образуют сплошной липидный бислой. Молекулы белков не образуют сплошного слоя, они располагаются в слое липидов, погружаясь в него на разную глубину. Молекулы белков и липидов подвижны.
12. Гиалоплазма, строение и функции. Цитоплазма, ее структурные компоненты.
Цитоплазма – это обязательный компонент клетки. Она размещена между плазмолеммой и кариолеммой. Структурными компонентами цитоплазмы является гиалоплазма, органеллы и включения.
Гиалоплазма (от греч. hyalinos – прозрачный), или матрикс цитоплазмы, представляет собой очень важную часть клетки, ее внутренняя среда. Гиалоплазма – наиболее жидкая часть цитоплазмы, в которой содержатся органеллы и включения. В общем объеме цитоплазмы гиалоплазма составляет около 50%. Она включает цитозоль (воду с растворенными в ней неорганическими и органическими веществами) и цитоматрикс (трабекулярную сетку волокон белковой природы толщиной 2-3 нм).
Гиалоплазма является сложной коллоидной системой, включающей в себя различные биополимеры: белки, нуклеиновые кислоты, полисахариды и др.. Эта система способна переходить из жидкого состояния в гелеобразный и наоборот. В организованной, упорядоченной, многокомпонентной системе гиалоплазмы отдельные зоны могут менять свое агрегатное состояние в зависимости от условий или от функциональной задачи; в бесструктурной, на первый взгляд, гиалоплазме могут возникать и распадаться различные фибриллярные, нитчатые комплексы белковых молекул. В состав гиалоплазмы входят, главным образом, различные глобулярные белки. Они составляют 20-25% общего содержания белков в эукариотической клетке.
К важнейшим ферментам гиалоплазмы относятся ферменты метаболизма сахаров, азотистых оснований, аминокислот, липидов и других важных соединений, так же находятся ферменты активации аминокислот при синтезе белков, транспортные (трансферные) РНК (тРНК). В гиалоплазме с участием рибосом и полирибосом (полисом) происходит синтез белков, необходимых для собственно клеточных потребностей, для поддержания и обеспечения жизни данной клетки. Осмотические и буферные свойства клетки, в значительной степени определяются составом и структурой гиалоплазмы.
Важнейшая роль гиалоплазмы заключается в том, что это полужидкая среда объединяет все клеточные структуры и обеспечивает химическое взаимодействие их друг с другом. Через гиалоплазму осуществляется большая часть внутриклеточных транспортных процессов: перенос аминокислот, жирных кислот, нуклеотидов, сахаров. В гиалоплазме идет постоянный поток ионов к плазматической мембране и от нее к митохондриям, к ядру и вакуолей. Гиалоплазма является основным вместилищем и зоной перемещения массы молекул АТФ. В гиалоплазме происходит отложение запасных продуктов: гликогена, жировых капель некоторых пигментов.
Строение и функции оболочки клетки. — КиберПедия
Клетка любого организма, представляет собой целостную живую систему. Она состоит из трех неразрывно связанных между собой частей: оболочки, цитоплазмы и ядра. Оболочка клетка осуществляет непосредственное взаимодействие с внешней средой и взаимодействие с соседними клетками (в многоклеточных организмах).
Оболочка клеток. Оболочка клеток имеет сложное строение. Она состоит из наружного слоя и расположенной под ним плазматической мембраны. Клетки животных и растений различаются по строению их наружного слоя. У растений, а также у бактерий, сине-зеленых водорослей и грибов на поверхности клеток расположена плотная оболочка, или клеточная стенка. У большинства растений она состоит из клетчатки. Клеточная стенка играет исключительно важную роль: она представляет собой внешний каркас, защитную оболочку, обеспечивает тургор растительных клеток: через клеточную стенку проходит вода, соли, молекулы многих органических веществ.
Наружный слой поверхности клеток животных в отличие от клеточных стенок растений очень тонкий, эластичный. Он не виден в световой микроскоп и состоит из разнообразных полисахаридов и белков.
Имея незначительную толщину (меньше 1 мкм), наружный слой клетки животных не выполняет опорной роли, какая свойственна клеточным стенкам растений.
Плазматическая мембрана. Под оболочкой клеток растений расположена плазматическая мембрана (лат. "мембрана"-кожица, пленка), граничащая непосредственно с цитоплазмой. Толщина плазматической мембраны около 10 нм, изучение ее строения и функций возможно только с помощью электронного микроскопа. В состав плазматической мембраны входят белки и липиды. Плазматическая мембрана выполняет много важных функций, от которых зависят жизнедеятельность клеток. Одна из таких функций заключается в том, что она образует барьер, отграничивающий внутреннее содержимое клетки от внешней среды. Но между клетками и внешней средой постоянно происходит обмен веществ. Из внешней среды в клетку поступает вода, разнообразные соли в форме отдельных ионов, неорганические и органические молекулы. Они проникают в клетку через очень тонкие каналы плазматической мембраны. Во внешнюю среду выводятся продукты, образованные в клетке. Транспорт веществ- одна из главных функций плазматической мембраны. Через плазматическую мембрану из клети выводятся продукты обмена, а также вещества, синтезированные в клетке. К числу их относятся разнообразные белки, углеводы, гормоны, которые вырабатываются в клетках различных желез и выводятся во внеклеточную среду в форме мелких капель.
На поверхности многих клеток животных, например, различных эпителиев, находятся очень мелкие тонкие выросты цитоплазмы, покрытые плазматической мембраной, - микроворсинки. Наибольшее количество микроворсинок находится на поверхности клеток кишечника, где происходит интенсивное переваривание и всасывание переваренной пищи.
Фагоцитоз. Крупные молекулы органических веществ, например белков и полисахаридов, частицы пищи, бактерии поступают в клетку путем фагоцита (греч. "фагео" - пожирать). В фагоците непосредственное участие принимает плазматическая мембрана. В том месте, где поверхность клетки соприкасается с частицей какого-либо плотного вещества, мембрана прогибается, образует углубление и окружает частицу, которая в "мембранной упаковке" погружается внутрь клетки. Образуется пищеварительная вакуоль и в ней перевариваются поступившие в клетку органические вещества.
Цитоплазма. Отграниченная от внешней среды плазматической мембраной, цитоплазма представляет собой внутреннюю полужидкую среду клеток. В цитоплазму эукариотических клеток располагаются ядро и различные органоиды. Ядро располагается в центральной части цитоплазмы. В ней сосредоточены и разнообразные включения - продукты клеточной деятельности, вакуоли, а также мельчайшие трубочки и нити, образующие скелет клетки. В составе основного вещества цитоплазмы преобладают белки. В цитоплазме протекают основные процессы обмена веществ, она объединяет в одно целое ядро и все органоиды, обеспечивает их взаимодействие, деятельность клетки как единой целостной живой системы.
Эндоплазматическая сеть. Вся внутренняя зона цитоплазмы заполнена многочисленными мелкими каналами и полостями, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной. Эти каналы ветвятся, соединяются друг с другом и образуют сеть, получившую название эндоплазматической сети.
Эндоплазматическая сеть неоднородна по своему строению. Известны два ее типа - гранулярная и гладкая. На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец - рибосом, которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности.
Эндоплазматическая сеть выполняет много разнообразных функций. Основная функция гранулярной эндоплазматической сети - участие в синтезе белка, который осуществляется в рибосомах.
На мембранах гладкой эндоплазматической сети происходит синтез липидов и углеводов. Все эти продукты синтеза накапливаются н каналах и полостях, а затем транспортируются к различным органоидам клетки, где потребляются или накапливаются в цитоплазме в качестве клеточных включений. Эндоплазматическая сеть связывает между собой основные органоиды клетки.
Рибосомы. Рибосомы обнаружены в клетках всех организмов. Это микроскопические тельца округлой формы диаметром 15-20 нм. Каждая рибосома состоит из двух неодинаковых по размерам частиц, малой и большой.
В одной клетке содержится много тысяч рибосом, они располагаются либо на мембранах гранулярной эндоплазматической сети, либо свободно лежат в цитоплазме. В состав рибосом входят белки и РНК. Функция рибосом - это синтез белка. Синтез белка - сложный процесс, который осуществляется не одной рибосомой, а целой группой, включающей до нескольких десятков объединенных рибосом. Такую группу рибосом называют полисомой. Синтезированные белки сначала накапливаются в каналах и полостях эндоплазматической сети, а затем транспортируются к органоидам и участкам клетки, где они потребляются. Эндоплазматическая сеть и рибосомы, расположенные на ее мембранах, представляют собой единый аппарат биосинтеза и транспортировки белков.
Митохондрии. В цитоплазме большинства клеток животных и растений содержатся мелкие тельца (0,2-7 мкм) - митохондрии (греч. "митос" - нить, "хондрион" - зерно, гранула).
Митохондрии хорошо видны в световой микроскоп, с помощью которого можно рассмотреть их форму, расположение, сосчитать количество. Внутреннее строение митохондрий изучено с помощью электронного микроскопа. Оболочка митохондрии состоит из двух мембран - наружной и внутренней. Наружная мембрана гладкая, она не образует никаких складок и выростов. Внутренняя мембрана, напротив, образует многочисленные складки, которые направлены в полость митохондрии. Складки внутренней мембраны называют кристами (лат. "криста" - гребень, вырост) Число крист неодинаково в митохондриях разных клеток. Их может быть от нескольких десятков до нескольких сотен, причем особенно много крист в митохондриях активно функционирующих клеток, например мышечных.
Митохондрии называют "силовыми станциями" клеток, так как их основная функция - синтез аденозинтрифосфорной кислоты (АТФ). Эта кислота синтезируется в митохондриях клеток всех организмов и представляет собой универсальный источник энергии, необходимый для осуществления процессов жизнедеятельности клетки и целого организма.
Новые митохондрии образуются делением уже существующих в клетке митохондрий.
Пластиды. В цитоплазме клеток всех растений находятся пластиды. В клетках животных пластиды отсутствуют. Различают три основных типа пластид: зеленые - хлоропласты; красные, оранжевые и желтые - хромопласты; бесцветные - лейкопласты.
Хлоропласт. Эти органоиды содержатся в клетках листьев и других зеленых органов растений, а также у разнообразных водорослей. Размеры хлоропластов 4-6 мкм, наиболее часто они имеют овальную форму. У высших растений в одной клетке обычно бывает несколько десятков хлоропластов. Зеленый цвет хлоропластов зависит от содержания в них пигмента хлорофилла. Xлоропласт - основной органоид клеток растений, в котором происходит фотосинтез, т. е. образование органических веществ (углеводов) из неорганических (СО2 и Н2О) при использовании энергии солнечного света.
В хлоропластах синтезируется и АТФ. Между внутренними мембранами хлоропласта содержатся ДНК, РНК. и рибосомы. Следовательно, в хлоропластах, так же как и в митохондриях, происходит синтез белка, необходимого для деятельности этих органоидов. Хлоропласты размножаются делением.
Хромопласты находятся в цитоплазме клеток разных частей растений: в цветках, плодах, стеблях, листьях. Присутствием хромопластов объясняется желтая, оранжевая и красная окраска венчиков цветков, плодов, осенних листьев.
Лейкопласты находятся в цитоплазме клеток неокрашенных частей растений, например в стеблях, корнях, клубнях. Форма лейкопластов разнообразна.
Хлоропласты, хромопласты и лейкопласты способны к взаимному переходу. Так при созревании плодов или изменении окраски листьев осенью хлоропласты превращаются в хромопласты, а лейкопласты могут превращаться в хлоропласты, например, при позеленении клубней картофеля.
Аппарат Гольджи. Во многих клетках животных, например в нервных, он имеет форму сложной сети, расположенной вокруг ядра. В клетках растений и простейших аппарат Гольджи представлен отдельными
Аппарат Гольджи выполняет много важных функций. Например, в клетках поджелудочной железы млекопитающих синтезируются пищеварительные ферменты, которые накапливаются в полостях органоида. На его мембранах происходит синтез жиров и углеводов (полисахаридов), которые используются в клетке и которые входят в состав мембран. Благодаря деятельности аппарата Гольджи происходят обновление и рост плазматической мембраны.
Лизосомы. Представляют собой небольшие округлые тельца. От цитоплазмы каждая лизосома отграничена мембраной. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты.
Обладая способностью к активному перевариванию пищевых веществ, лизосомы участвуют в удалении отмирающих в процессе жизнедеятельности частей клеток, целых клеток и органов. Образование новых лизосом происходит в клетке постоянно. Ферменты, содержащиеся в лизосомах, как и всякие другие белки синтезируются на рибосомах цитоплазмы. Затем эти ферменты поступают по каналам эндоплазматической сети к аппарату Гольджи, в полостях которого формируются лизосомы. В таком виде лизосомы поступают в цитоплазму.
Клеточный центр. В клетках животных вблизи ядра находится органоид, который называют клеточным центром. Основную часть клеточного центра составляют два маленьких тельца - центриоли, расположенные в небольшом участке уплотненной цитоплазмы играют важную роль при делении клетки; они участвуют в образовании веретена деления.
Клеточные включения. К клеточным включениям относятся углеводы, жиры и белки. Все эти вещества накапливаются в цитоплазме клетки в виде капель и зерен различной величины и формы. Они периодически синтезируются в клетке и используются в процессе обмена веществ.
Ядро. Каждая клетка одноклеточных и многоклеточных животных, а также растений содержит ядро. Форма и размеры ядра зависят от формы и размера клеток. В большинстве клеток имеется одно ядро, и такие клетки называют одноядерными. Существуют также клетки с двумя, тремя, с несколькими десятками и даже сотнями ядер. Это - многоядерные клетки.
Ядерный сок - полужидкое вещество, которое находится под ядерной оболочкой и представляет внутреннюю среду ядра.
Клеточное ядро — Википедия
У этого термина существуют и другие значения, см. Ядро. Клетки HeLa, ДНК окрашена синим красителем Hoechst[en]. Центральная и правая клетка находятся в интерфазе, и у них окрашено всё ядро. Левая клетка претерпевает митоз, поэтому ДНК конденсирована и окрашено не всё ядро
Клетки HeLa, ДНК окрашена синим красителем Hoechst[en]. Центральная и правая клетка находятся в интерфазе, и у них окрашено всё ядро. Левая клетка претерпевает митоз, поэтому ДНК конденсирована и окрашено не всё ядро Кле́точное ядро́ (лат. nucleus) — окружённый двумя мембранами компартмент эукариотической клетки[1] (в клетках прокариот ядро отсутствует). Обычно в клетках эукариот имеется одно ядро, однако некоторые типы клеток, например, эритроциты млекопитающих, не имеют ядра, а другие содержат несколько ядер.
В ядре заключена бо́льшая часть генетического материала клетки, представленного несколькими линейными длинными молекулами ДНК, связанного с белками — хромосомами. Гены, локализованные в хромосомах, составляют ядерный геном. Ядро поддерживает целостность генов, а входящие в его состав белки регулируют клеточные процессы посредством управления экспрессией генов, поэтому ядро является, по сути, контролирующим центром клетки. К основным структурам, из которых состоит ядро, относят ядерную оболочку — двойную мембрану, окружающую ядро и изолирующую его от цитоплазмы, а также ядерный матрикс (который включает ядерную ламину) — сеть филаментов, которая обеспечивает механическую поддержку ядра, подобно цитоскелету в цитоплазме.
Поскольку ядерная оболочка непроницаема для крупных молекул, для регуляции транспорта молекул через ядерную оболочку (ядерный транспорт[en]) служат ядерные поры. Поры пронизывают обе ядерные мембраны и формируют сквозной канал, через который малые молекулы и ионы проходят свободно, а крупные молекулы активно транспортируются с участием белков-переносчиков. Перенос через ядерную оболочку таких крупных молекул, как белки и РНК, необходим для экспрессии генов и поддержания хромосом. Хотя внутри ядра нет окружённых мембраной субкомпартментов, его внутреннее содержимое неоднородно и содержит ряд ядерных телец, которые состоят из особых белков, молекул РНК и частей хромосом. Самое известное ядерное тельце — ядрышко, в котором происходит сборка рибосомных субъединиц. После образования в ядрышке рибосомные субъединицы транспортируются в цитоплазму, где они осуществляют трансляцию мРНК.
Ядро стало первой из органелл, открытых учёными-естествоиспытателями в составе клетки. Самые ранние рисунки клеток и их ядер принадлежат основоположнику научной микроскопии Антони ван Левенгуку (1633—1723), который наблюдал ядро в эритроцитах лосося[2]. Описания ядра также выполнил Франц Бауэр[en] в 1802 году[3], а более детальное описание было выполнено в 1831 году шотландским ботаником Робертом Броуном и представлено на собрании Лондонского Линнеевского общества. Броун изучал орхидеи под микроскопом и обнаружил в клетках наружного слоя цветка непрозрачные области, которые он называл «ареолами» или «ядрами»[4].
Броун не делал предположений относительно функций ядра. В 1838 году Маттиас Шлейден предположил, что ядро участвует в образовании новых клеток, поэтому он ввёл для обозначения ядер термин «цитобласт» (клеточный строитель). Он был уверен, что наблюдал сборку новых клеток вокруг «цитобластов». Убеждённым оппонентом этого взгляда был Франц Мейен, открывший, что клетки размножаются посредством деления, и считавший, что у многих клеток может не быть ядра. Идея об образовании клеток de novo[en], то есть с нуля, посредством цитобластов или иначе, противоречила работам Роберта Ремака (1852) и Рудольфа Вирхова (1855), которые окончательно утвердили новую парадигму, утверждающую, что клетки могут образовываться только из клеток («Omnis cellula e cellula»). Функции ядра оставались неясными[5].
Между 1877 и 1878 годами Оскар Гертвиг опубликовал несколько работ по оплодотворению яиц у морских ежей, в которых показал, что при оплодотворении ядро сперматозоида проникает внутрь яйцеклетки и сливается с её ядром. Впервые было показано, что новая особь развивается из единственной клетки, имеющей ядро. Это противоречило теории Эрнста Геккеля, согласно которой в ходе эмбрионального развития особи последовательно проходятся все этапы филогении её вида, а потому, в частности, поколение первых клеток с ядром якобы образуется из «монерулы» — бесструктурной массы первичной слизи. В связи с этим необходимость ядра сперматозоида для оплодотворения некоторое время была предметом дискуссий. Однако Гертвиг подтвердил свои наблюдения исследованиями на других животных, включая земноводных и моллюсков. В 1884 году Эдуард Страсбургер показал то же самое для растений. Это проложило путь к гипотезе о том, что ядро передаёт наследственный материал. В 1873 году Август Вейсман высказал идею о равнозначности материнского и отцовского материала для наследственности. Функция ядра как носителя генетической информации стала очевидной лишь позже, после открытия митоза и открытия заново законов Менделя в начале XX столетия. На основании этих открытий была сформулирована хромосомная теория наследственности[5].
Ядро — крупнейшая органелла животных клеток[6]. У млекопитающих диаметр ядра составляет примерно 6 мкм, а само ядро составляет около 10 % объёма клетки[7]. Вязкая жидкость, заполняющая ядро, называется нуклеоплазмой и по химическому составу близка к цитозолю, окружающему ядро[8].
Ядерная оболочка и ядерные поры[править | править код]
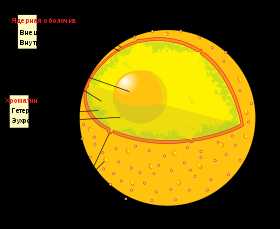 Строение клеточного ядра Строение клеточного ядра | 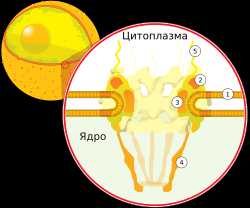 Поперечный разрез ядерной поры. Цифрами обозначены: 1 — ядерная оболочка, 2 — внешнее кольцо, 3 — спицы, 4 — корзина, 5 — филаменты Поперечный разрез ядерной поры. Цифрами обозначены: 1 — ядерная оболочка, 2 — внешнее кольцо, 3 — спицы, 4 — корзина, 5 — филаменты |
Ядерная оболочка состоит из двух мембран (наружной и внутренней), которые расположены параллельно на расстоянии от 10 до 50 нм. Ядерная оболочка полностью окружает ядро, отделяя генетический материал клетки от цитоплазмы и служа барьером, предотвращающим свободную диффузию макромолекул между нуклеоплазмой и цитоплазмой. Наружная ядерная мембрана продолжается в мембрану шероховатого эндоплазматического ретикулума (ЭПР) и покрыта рибосомами. Промежуток между ядерными мембранами называется перинуклеарным пространством и продолжается в люмен ЭПР[9].
Ядерные поры, представляющие собой заполненные водой каналы в ядерной оболочке[1], состоят из множества белков, называемых нуклеопоринами. У человека масса пор составляет около 120 000 кДа,что в 40 раз больше массы рибосомы[10]; при этом у дрожжей в состав ядерных пор входит около 50 белков, а у позвоночных — несколько сотен[6]. Хотя диаметр пор составляет 100 нм, ширина щели, через которую могут проходить молекулы, из-за наличия внутри пор регуляторных систем составляет всего 9 нм. В такую щель могут проходить водорастворимые малые молекулы, но не крупные молекулы — такие, как нуклеиновые кислоты и большие белки; для переноса этих молекул в ядро необходим активный (то есть энергозатратный) транспорт. На оболочке ядра типичной клетки млекопитающего располагается от 3000 до 4000 пор[11], и у каждой в месте слияния двух ядерных мембран находится кольцевая структура, имеющая 8 осей симметрии[12]. К кольцу прикрепляется особая структура, известная как ядерная корзина, которая выдаётся в нуклеоплазму, а несколько её филаментов выдаются в цитоплазму. Обе структуры необходимы для опосредования связывания транспортных ядерных белков[6].
Большинство белков, субъединицы рибосом и некоторые ДНК переносятся через ядерные поры посредством семейства транспортных факторов, известных как кариоферины[en]. Кариоферины, опосредующие транспорт в ядро, также называются импортинами[en], а опосредующие транспорт из ядра — экспортинами. Большинство кариоферинов непосредственно взаимодействуют со своим грузом, но некоторые используют для этого адаптерные[en] белки[13]. Стероидные гормоны (такие, как кортизол и альдостерон), а также другие жирорастворимые малые молекулы могут диффундировать в цитоплазму внутрь клетки через клеточную мембрану; в цитоплазме они связываются с белковыми ядерными рецепторами, которые доставляют их в ядро. Здесь ядерные рецепторы[en], связанные со своими лигандами, функционируют как транскрипционные факторы, а в отсутствие лиганда многие рецепторы функционируют как гистондеацетилазы, подавляющие экспрессию некоторых генов[6].
Ядерная ламина[править | править код]
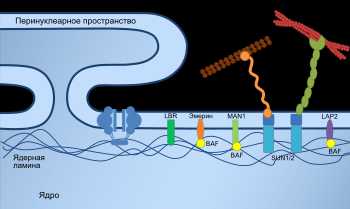 Строение ядерной оболочки и ядерной ламины
Строение ядерной оболочки и ядерной ламины В клетках животных механическую поддержку ядра обеспечивают две сети из промежуточных филаментов: ядерная ламина, представляющая собой сеть промежуточных филаментов на внутренней поверхности ядра, а также менее организованные филаменты на цитозольной поверхности ядра. Обе системы филаментов обеспечивают поддержку ядра и служат для закрепления хромосом и ядерных пор[7].
Ядерная ламина состоит в основном из белков, известных как ламины. Как и все белки, ламины синтезируются в цитоплазме и далее транспортируются внутрь ядра, где они вставляются в ядерную ламину[14][15]. Расположенные на наружной стороне ядерной оболочки ламины (такие, как эмерин[en] и несприн[en]), связываются с элементами цитоскелета, что обеспечивает структурную поддержку ядру. Ламины также обнаруживаются в нуклеоплазме, где они образуют другую регулярную структуру, известную как нуклеоплазматическая вуаль (англ. nucleoplasmic veil)[16]; последнюю можно визуализировать с использованием флуоресцентной микроскопии. Функция вуали неизвестна, но известно, что её нет в ядрышке и она присутствует в интерфазе клеточного цикла[17]. Входящие в состав вуали ламины (такие, как LEM3) связываются с хроматином, и нарушения в их структуре подавляют транскрипцию белоккодирующих генов[18].
Как и другие белки промежуточных филаментов, мономеры ламинов содержат α-спиральный домен, используемый двумя мономерами, чтобы обвиться вокруг друг друга, образуя димер, имеющий структуру биспирали[en]. Два димера далее связываются своими боковыми сторонами в антипараллельной ориентации, образуя тетрамер, известный как протофиламент. Восемь тетрамеров объединяются в скрученный, похожий на верёвку филамент. Филаменты могут собираться и разбираться динамическим образом, то есть длина филамента зависит от относительных скоростей его сборки и разборки[7].
Хромосомы[править | править код]
 Хромосомные территории 24 хромосом человека
Хромосомные территории 24 хромосом человека В ядре находится большая часть генетического материала клетки, представленного множеством линейных молекул ДНК, которые организованы в структуры, известные как хромосомы. Суммарная длина молекул ДНК клетки человека составляет около 2 м. В течение большей части клеточного цикла данные молекулы в комплексе с белками формируют так называемый хроматин, а при клеточном делении хромосомы предстают в виде отдельных хорошо различимых хромосом, составляющих кариотип. Небольшое количество клеточного генетического материала располагается в митохондриях и, в случае растительной клетки, в хлоропластах[19].
Известно два вида хроматина. В эухроматине ДНК наименее плотно организована; он содержит гены, которые транскрибируются наиболее часто[19]. Другой вид хроматина, гетерохроматин, более компактен и содержит ДНК, транскрибируемую редко или никогда. Гетерохроматин подразделяется на факультативный, который присутствует только в клетках определённого типа и на определённой стадии клеточного цикла, и конститутивный, представленный такими структурами хромосом, как теломеры и центромеры[20]. В течение интерфазы хроматин каждой хромосомы занимает строго определённую область ядра — хромосомную территорию[21][22]. Активные гены, которые, как правило, располагаются в эухроматине, обычно располагаются на границе хромосомной территории[23].
Ядерные тельца[править | править код]
В ядре клеток млекопитающих содержится ряд дискретных субкомпартментов[24], которые называются ядерными тельцами. Они осуществляют компартментализацию ядра, создавая внутри него отдельные пространства, которые обладают определёнными свойствами. Многие ядерные тельца осуществляют специфические функции — например, синтез и процессинг пре-рибосомных РНК в ядрышке, накопление и сборку компонентов сплайсосом в спеклах (см. ниже) или накопление молекул РНК в параспеклах. Механизмы, которые обеспечивают выполнение ядрышковыми тельцами этих функций, очень разнообразны. В некоторых случаях ядерное тельце может служить местом протекания определённых процессов, например, транскрипции. В других случаях ядерные тельца, по-видимому, опосредованно регулируют локальные концентрации своих компонентов в нуклеоплазме. Подобно цитоплазматическим органеллам, ядерные тельца содержат специфический набор белков, которые определяют их структуру на молекулярном уровне. Однако, в отличие от органелл цитоплазмы, ядерные тельца не окружены липидными мембранами, и их структурная целостность целиком обеспечивается белок-белковыми и РНК-белковыми взаимодействиями. Ниже в таблице перечислены основные характеристики ядерных телец[25].
| Ядерное тельце | Функции | Характерные компоненты | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|---|
| Ядрышко | Биогенез рибосом | Машинерия РНК-полимеразы I[en], факторы процессинга рРНК и сборки рибосомных субъединиц | 3—8 | 1—4 |
| Спеклы | Накопление и сборка факторов сплайсинга | Факторы сплайсинга пре-мРНК | 2—3 | 20—50 |
| Стрессовые ядерные тельца | Регуляция транскрипции и сплайсинга в условиях стресса | HSF1[en], HAP | 1—2 | 3—6 |
| Тельце гистоновых локусов | Процессинг пре-мРНК гистонов | NPAT[en], FLASH, U7[en] мяРНП | 0,2—1,2 | 2—4 |
| Тельце Кахаля | Биогенез, созревание и кругооборот малых РНК | Коилин, SMN[en] | 0,2—1,5 | 1—10 |
| PML-тельце | Регуляция стабильности генома, репарация ДНК, контроль транскрипции, защита от вирусов | PML | 0,1—1 | 10—30 |
| Параспеклы | Регуляция мРНК, редактирование РНК | Некодирующие РНК NEAT1/MENε/β, белки PSP1, p54nrb/NONO | 0,2—1 | 2—20 |
| Околоядрышковый компартмент | Посттранскрипционная регуляция набора РНК, синтезированных РНК-полимеразой III[en] | PTB | 0,2—1 | 1—2 |
Ядрышко[править | править код]
Ядрышко — это отдельная плотная структура в ядре. Она не окружена мембраной и формируется в области расположения рДНК — тандемных повторов генов рибосомной РНК (рРНК), называемых ядрышковыми организаторами. Главная функция ядрышка — синтез рРНК и образование рибосом. Структурная целостность ядрышка зависит от его активности, и инактивация генов рРНК приводит к смешению ядрышковых структур[26].
На первой стадии образования рибосом фермент РНК-полимераза I транскрибирует рДНК и образует пре-рРНК, которая далее разрезается на 5,8S, 18S и 28S рРНК[27]. Транскрипция и посттранскрипционный процессинг рРНК происходят в ядрышке при участии малых ядрышковых РНК (snoРНК), некоторые из которых происходят из сплайсированных интронов мРНК генов, кодирующих белки, связанные с работой рибосом. Собранные рибосомные субъединицы — это самые крупные структуры, проходящие через ядерные поры[6].
При рассматривании под электронным микроскопом в ядрышке можно выделить три компонента: фибриллярные центры (ФЦ), окружающий их плотный фибриллярный компонент (ПФК) и гранулярный компонент (ГК), который, в свою очередь, окружает ПФК. Транскрипция рРНК происходит в ФЦ и на границе ФЦ и ПФК, поэтому при активации образования рибосом ФЦ становятся хорошо различимы. Разрезание и модификации рРНК происходят в ПФК, а последующие этапы образования рибосомных субъединиц, включающие загрузку рибосомных белков, происходят в ГК[27].
Тельце Кахаля[править | править код]
 Ядра клеток мыши (синие), содержащие тельца Кахаля (зелёные точки). Изображение получено методом флуоресцентной микроскопии (коилин — маркер телец Кахаля — сращён с зелёным флуоресцентным белком)
Ядра клеток мыши (синие), содержащие тельца Кахаля (зелёные точки). Изображение получено методом флуоресцентной микроскопии (коилин — маркер телец Кахаля — сращён с зелёным флуоресцентным белком) Тельце Кахаля (ТК) — ядерное тельце, имеющееся у всех эукариот. Оно идентифицируется по наличию сигнатурного белка коилина и специфических РНК (scaРНК). В ТК также содержится белок SMN (англ. survival of motor neurons). В ТК наблюдается высокая концентрация сплайсирующих малых ядерных рибонуклеопротеинов (мяРНП) и других факторов процессинга РНК, поэтому считается, что ТК служат местами сборки и/или посттранскрипционной модификации факторов сплайсинга. ТК присутствует в ядре во время интерфазы, но исчезает в митозе. В биогенезе ТК прослеживаются свойства самоорганизующейся структуры[28].
Когда внутриклеточная локализация SMN впервые изучалась методом иммунофлуоресценции, то белок обнаруживался во всей цитоплазме, а также в ядрышковом тельце, сходном по размеру с ТК и часто расположенном рядом с ТК. По этой причине данное тельце было названо «близнецом ТК» (англ. gemini of CB) или просто gem. Однако оказалось, что линия клеток HeLa, в которой было открыто новое тельце, была необычной: в других линиях клеток человека, а также у плодовой мушки Drosophila melanogaster SMN колокализовался с коилином в ТК. Поэтому в общем случае SMN можно рассматривать как важный компонент ТК, а не как маркер отдельного ядерного тельца[29].
Тельце гистоновых локусов[править | править код]
Тельце гистоновых локусов (англ. histone locus body, HLB) содержит факторы, необходимые для процессинга пре-мРНК гистонов. Как и следует из названия, тельца гистоновых локусов ассоциированы с генами, кодирующими гистоны; поэтому предполагается, что в тельцах гистоновых локусов концентрируются факторы сплайсинга. Тельце гистоновых локусов присутствует в клетке во время интерфазы и исчезает с наступлением митоза. Тельце гистоновых локусов нередко рассматривается вместе с тельцем Кахаля по нескольким причинам. Во-первых, в некоторых тельцах гистоновых локусов содержится маркер телец Кахаля — коилин. Во-вторых, эти тельца нередко физически находятся рядом, поэтому между ними наблюдается некоторое взаимодействие. Наконец, очень крупные тельца Кахаля ооцитов земноводных обладают свойствами обоих телец[28].
PML-тельца[править | править код]
Тельца промиелоцитной лейкемии (англ. Promyelocytic leukaemia bodies), или PML-тельца — сферические тельца, разбросанные по всей нуклеоплазме и достигающие около 0,1—1,0 мкм в диаметре. Они известны также под такими названиями, как ядерный домен 10 (англ. nuclear domain 10 (ND10)), тельца Кремера (англ. Kremer bodies) и онкогенные домены PML (англ. PML oncogenic domains). Тельца PML названы по одному из своих ключевых компонентов — белку промиелоцитной лейкемии (PML). Они часто наблюдаются ассоциированными с тельцами Кахаля и тельцами деления (англ. cleavage body)[30]. PML-тельца принадлежат ядерному матриксу и могут быть задействованы в таких процессах, как репликация ДНК, транскрипция и эпигенетический сайленсинг генов[31]. Ключевым фактором организации этих телец выступает белок PML, который привлекает другие белки; последние, по современным представлениям, объединены лишь тем, что они SUMOилированы[en]. Мыши, у которых ген PML делетирован, лишены PML-телец, однако развиваются и живут нормально, поэтому PML-тельца не выполняют незаменимых биологических функций[31].
Спеклы[править | править код]
Спеклы (англ. speckle) — это ядерные тельца, которые содержат факторы сплайсинга пре-мРНК и располагаются в интерхроматиновых участках нуклеоплазмы клеток млекопитающих. При флуоресцентной микроскопии спеклы выглядят как пятнистые тельца неправильной формы, различных размеров, а при электронной микроскопии они выглядят как кластеры интерхроматиновых гранул. Спеклы — динамические структуры, и содержащиеся в них белки и РНК могут перемещаться между спеклами и другими ядерными тельцами, включая участки активной транскрипции. На основании исследований состава, структуры и поведения спеклов была создана модель, объясняющая функциональную компартментализацию ядра и организацию машинерии экспрессии генов[32], сплайсирующих малые ядерные рибонуклеопротеины[33][34] и другие белки, необходимые для сплайсинга пре-мРНК[32]. Из-за изменяющихся потребностей клетки состав и расположение спеклов изменяется согласно транскрипции мРНК и посредством регуляции фосфорилирования специфических белков[35]. Сплайсирующие спеклы также известны как ядерные спеклы, компартменты сплайсирующих факторов, кластеры интерхроматиновых гранул и B-снурпосомы (англ. B snurposomes)[36]. B-снурпосомы найдены в ядрах ооцитов земноводных и зародышах плодовой мушки Drosophila melanogaster[37]. На электронных микрофотографиях B-снурпосомы предстают прикреплёнными к тельцам Кахаля или отдельно от них. Кластеры интерхроматиновых гранул служат местами скопления факторов сплайсинга[38].
Параспеклы[править | править код]
 Микрофотография клеток HeLa с меченым белком параспекл PSP1: 1. цитоплазма; 2. ядро; 3. ядрышко; 4. параспеклы
Микрофотография клеток HeLa с меченым белком параспекл PSP1: 1. цитоплазма; 2. ядро; 3. ядрышко; 4. параспеклы Параспеклы — это ядерные тельца неправильной формы, располагающиеся в интерхроматиновом пространстве ядра[39]. Впервые они были описаны у клеток HeLa, у которых имеется 10—30 параспеклов на ядро, но сейчас параспеклы обнаружены во всех первичных клетках человека, в клетках трансформированных линий и на срезах тканей[40]. Своё название они получили из-за своего расположения в ядре — вблизи спеклов[39].
Параспеклы — динамические структуры, которые изменяются в ответ на изменения в метаболической активности клетки. Они зависят от транскрипции[39], и в отсутствие транскрипции, проводимой РНК-полимеразой II, параспеклы исчезают, а все входящие в их состав белки (PSP1, p54nrb, PSP2, CFI(m)68 и PSF) формируют серповидный околоядрышковый кэп. Этот феномен наблюдается в ходе клеточного цикла: параспеклы присутствуют в интерфазе и всех фазах митоза, за исключением телофазы. В ходе телофазы формируются дочерние ядра, и РНК-полимераза II ничего не транскрибирует, поэтому белки параспеклов и формируют околоядрышковый кэп[40]. Параспеклы участвуют в регуляции экспрессии генов, накапливая те РНК, где есть двухцепочечные участки, которые подвергаются редактированию, а именно превращению аденозина в инозин. Благодаря этому механизму параспеклы задействованы в контроле экспрессии генов при дифференцировке, вирусной инфекции и стрессе[41].
Околоядрышковый компартмент[править | править код]
Околоядрышковый компартмент (ОК) — ядерное тельце неправильной формы, которое характеризуется тем, что располагается на периферии ядрышка. Несмотря на физическую связь, эти два компартмента структурно различны. Обычно ОК обнаруживают в клетках злокачественных опухолей[42]. ОК — динамическая структура, и содержит очень много РНК-связывающих белков и РНК-полимеразу III. Структурная стабильность ОК обеспечивается транскрипцией, осуществляемой РНК-полимеразой III, и наличием ключевых белков. Поскольку присутствие ОК обычно связано со злокачественностью и со способностью к метастазированию, их рассматривают как потенциальные маркеры рака и других злокачественных опухолей. Показана ассоциация ОК со специфическими локусами ДНК[43].
Стрессовые ядерные тельца[править | править код]
Стрессовые ядерные тельца формируются в ядре при тепловом шоке. Они образуются при непосредственном взаимодействии транскрипционного фактора теплового шока 1 (HSF1[en]) и перицентрических тандемных повторов в последовательности сателлита III, что соответствует сайтам активной траснкрипции некодирующих транскриптов сателлита III. Распространено мнение, что такие тельца соответствуют очень плотно упакованным формам рибонуклеопротеиновых комплексов. Считается, что в клетках, подвергающихся стрессу, они участвуют в быстрых, временных и глобальных изменениях в экспрессии генов посредством различных механизмов — например, ремоделирования хроматина и захватывания факторов транскрипции и сплайсинга. В клетках, находящихся в нормальных (не стрессовых) условиях, стрессовые ядерные тельца обнаруживаются редко, однако их количество резко увеличивается под действием теплового шока. Стрессовые ядерные тельца найдены только в клетках человека и других приматов[44].
Ядерные тельца-сироты[править | править код]
Ядерные тельца-сироты (англ. orphan nuclear bodies) — нехроматиновые ядерные компартменты, которые исследованы гораздо хуже, чем другие хорошо охарактеризованные структуры ядра. Некоторые из них выступают как места, в которых белки модифицируются белками SUMO и/или происходит протеасомная деградация белков, помеченных убиквитином[45]. Ниже в таблице приведены характеристики известных ядерных телец-сирот[46].
| Ядерное тельце | Описание | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|
| Кластосома | Концентрирует протеасомные комплексы 20S и 19S и белки, связанные с убиквитином. Обнаруживается, главным образом, тогда, когда стимулируется активность протеасом, и разбирается при ингибировании активности протеасом. | 0,2—1,2 | 0—3 |
| Тельце деления (англ. cleavage body) | Обогащено факторами деления CstF[en] и CPSF[en], а также белком DDX1[en], содержащим DEAD-бокс[en]. Обнаруживается в основном в S-фазе, ингибирование транскрипции на него не влияет. | 0,2—1,0 | 1—4 |
| Домен OPT | Обогащён факторами транскрипции Oct1[en] и PTF. Частично колокализуется с сайтами транскрипции. Обнаруживается в основном в поздней G1-фазе, разбирается при ингибировании транскрипции. | 1,0—1,5 | 1—3 |
| Тельце Polycomb | Обнаруживается в клетках человека и дрозофилы, обогащено белком PcG. У человека накапливает белки RING1, BMI1[en], HPC, может быть связано с околоцентромерным гетерохроматином. | 0,3—1,0 | 12—16 |
| Тельце Sam68 | Накапливает белок Sam68 и схожие с ним белки SLM-1 и SLM-2. Разбирается при ингибировании транскрипции. Вероятно, обогащено РНК. | 0,6—1,0 | 2—5 |
| Тельце SUMO | Обогащено белками SUMO и SUMO-конъюгирующим ферментом Ubc9[en]. Концентрирует транскрипционные факторы pCREB, CBP, c-Jun[en]. | 1—3 | 1—3 |
Ядерная оболочка защищает ДНК клетки и участвует в гораздо более сложной регуляции экспрессии генов по сравнению с прокариотической клеткой. У прокариот транскрипция и трансляция являются сопряжёнными процессами и трансляция мРНК в белок начинается ещё до того, как она будет полностью синтезирована. В клетках эукариот цитоплазма, в которой проходит трансляция, и транскрипция, протекающая в ядре, пространственно разобщены, поэтому возникает необходимость в обеспечени