Неспецифические факторы иммунитета
70. Понятие иммунитета. Неспецифические факторы защиты
Иммунитет – способность поддерживать генеостаз (постоянство внутренней среды). Все средства защиты разделяются на специфические и неспецифические. Специфические - появляется иммунитет через 48 часов после контакта с патогенном (латентный период) и действует против строго определённого патогенна – адентивный иммунный ответ. Неспецифические факторы защиты препятствуют размножению патогена в латентный период; действуют против любого патогенна с разной эффективностью – воспалительная реакция. К ним относят кожа и слизистая, клеточная защита, гуморальная защита. Кожа и слизистая: явл-ся преградой на пути микробов; на поверхности кожи высокое осмотическое давление, молочная кислота, ненасыщенные жирные кислоты; слизистая выделяет секреты с бактерицидными свойствами, в том числе желудочный и кишечный сок. Всё это неблагоприятно для развития микробов. Если микробы внедрились в подкожную клетчатку или в подслизистую ткань, место внедрения – ворота инфекции. Сюда устремляется группа фагоцитов. Клеточная защита – фагоцитоз. Гл роль в нем играют лейкоциты – при остром воспалении; фагоциты – при хроническом воспалении. Микроб, захваченный фагоцитом, может подвергнуться полному перевариванию – завершённый фагоцитоз. Микроб внутри лейкоцита размножается – незавершённый фагоцитоз. В таком виде патоген не доступен действию антител. Многие микробы имеют капсулы, выделяют токсины => полиинфекции покрываются погибшими эритроцитами микробов и продуктами распада. Развивается воспаление. В этот очаг поступает жидкая часть крови и лимфы, кот содерж гуморальные факторы защиты – стволовые лимфоидные клетки превращаются в В – лимфоциты, кот ответственны за реализацию гуморального иммунного ответа. В – система ответственна за иммунитет при многих бактериальных инфекциях, антитоксический иммунитет, аллергию немедленного типа. В – лимфоциты имеют рецепторы – макромолекулярные структуры клеточной поверхности, с помощью кот клетки узнают антигены.
72. Лимфоциты: т и в – типа. Их функции
Лимфоциты содержат крупное ядро, окружённым узким ободком слабо базафильной цитоплазмы, органоиды слабо развиты. По функционному признаку различают: Т – лимфоциты проходят развитие в вилочковой железе и в спец зонах переферич лимфоидных органоидов. Долгоживущие. Обеспечивают реакции клеточного иммунитета, участвуют в гуморальном иммунитете. Среди них различают: Т – клетки памяти – долго живут, сохраняя информацию об антигене, кот вызвал их появление. Т –киллеры – обладают цитотоксичным эффектом по отношению к чужеродным клеткам. Т- хелперы – помощники – в гуморальном иммунитете – помогают В- лимфоцитам вырабатывать иммуноглобулин. Т – супрессоры – подавляют способность В - лимфоцитов вырабатывать иммуноглобулин. В-лимфоциты развиваются в красном костном мозге и переферич лимфоидных органах. Короткоживущие. В – клетки памяти – сохраняют информацию об антигене.
73. Иммунный ответ. Локус иммунного ответа
Иммунный ответ, или иммунологическая реактивность, - высокоспецифическая форма реакции организма на чужеродные вещества (антигены). При иммунном ответе происходят распознавание чужеродного агента. При введении антигена возникает первичный иммунный ответ - через 2 дня в крови образуются антитела, титр которых возрастает, достигает максимума, а затем падает. Вторичный иммунный ответ возникает на повторное введение того же антигена и характеризуется более высоким и быстрым нарастанием титра антител. Подобная реакция более усиленного образования антител на повторное введение антигена – иммунологическая память При вирусной инфекции ДНК или РНК вируса попадает в клетку, а вирусные белки остаются на клеточной мембране. Цитотоксические Т-киллеры своими рецепторами узнают вирусные антигены только в комбинации с белком главного комплекса гистосовместимости МНС класса 1.В отличие от антител Т-рецепторы не узнают и не связывают антиген, если тот не находится вместе с белком МНС. После узнавания антигенов цитотоксические Т-клетки убивают зараженные вирусом клетки. Мутации любых локусов, обусловливающие разные звенья иммунной системы организма, влияют на иммунный ответ. Гены иммунного ответа. Гены, кодирующие иммунный ответ, наз-ся генами иммунного ответа Высота иммунного ответа детерминирована многими генами иммунного ответа, обозначаемыми Iг-1, Iг-2 и т. д. Контроль иммунного ответа осуществляется Iг-генами путем контроля синтеза Iа-белков. Во многих случаях иммунный ответ против антигенов наследуется полигенно. Гены иммунного ответа: 1) Ir-гены определяют количество синтезируемых антител против определенных антигенов; 2) Ir-гены не сцеплены с локусами, кодирующими иммуноглобулины; 3) Ir-гены высокоспецифичны. 4) между генами, контролирующими высокий или низкий иммунный ответ против различных антигенов, в основном не существует никакой связи. Теории иммунитета: 1) клонально-селекционная теория Ф. Бернета (1959). Она основана на четырех основных принципах: а) в организме имеется большое число лимфоидных клеток; б) популяция лимфоидных клеток гетерогенна, и в результате интенсивного деления клеток образуется большое число клонов; в) небольшое количество антигена стимулирует клон клеток к размножению; г) большое количество антигена элиминирует соответствующий клон. 2) Сетевая теория. Согласно неё антитела не только узнают антиген, но и сами являются антигенами.
Специфический и неспецифический иммунитет
Доброго времени суток, дорогие читатели!Продолжаем рассуждения не тему иммунитета и всего, что с ним связано. Сегодняшняя статья позволит углубиться нам ещё дальше в познания, связанные с защитными свойствами человека. С надежной охраной от биологических тел или веществ, несущих чужеродную генетическую информацию. Защитой, которая носит название иммунная система. Такие понятия как специфический и неспецифический иммунитет, раскроют перед нами тайны лейкоцитов, стоящих на страже нашего здоровья.
Содержание статьи:
Основные виды иммунитета
Защитная система человека представлена видовым разнообразием лейкоцитов, а также внутренними воспроизводителями по их выработке и развитию. Важные для нас лимфоциты, макрофаги, нейтрофилы, моноциты имеют происхождение из тканей костного мозга, селезенки, лимфоузлов и тимуса.
Целая вселенная, макрокосмос находится внутри нас. Все эти составляющие стоят на страже нашего здоровья, ежеминутно преобразовываясь и переходя из одного состояния в другое. Быстро и эффективно подавляя антигены, вещества чуждые человеческой природе. Среди видов иммунитета, направляющих свое действие на чужеродные микробы и бактерии, определяют неспецифический и специфический виды.
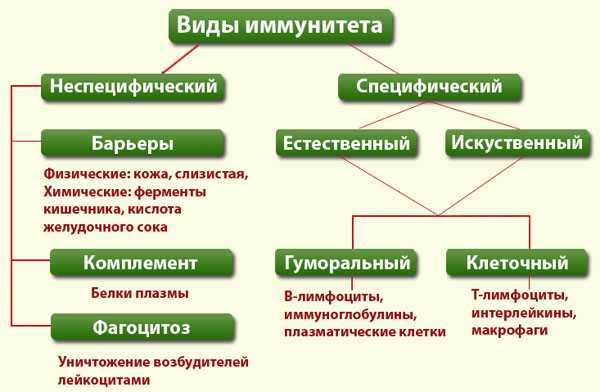
Неспецифический иммунитет
Данный тип, призван к борьбе с антигеном и проявляется гуморальным и клеточным видами. Первый работает в плазме крови и получается путём выработки бактерицидных составляющих, а клеточный говорит сам за себя, эта работа ведётся на клеточном уровне.
Неспецифический иммунитет ещё называют врожденным, видовым, он способен заблокировать любое посягательство на организм с внешней стороны. Воспринимает вторжение, как чужеродное и опасное, для внутренних систем человека.
Действует этот вид защиты влиянием на антиген различных видов лейкоцитов, в их числе нейтрофилы, макрофаги, базофилы. Это в общих чертах. Теперь несколько подробнее. Что такое неспецифический иммунитет?
Данный вид иммунитета имеет устойчивость к ряду вирусных инфекций. Будущий человек, находясь в материнской утробе, надёжно спрятан от антигена, даже после рождения, влияние наследственного иммунитета остается с ним на протяжении шести месяцев. Но, стоит заметить, что данный срок относится далеко не ко всем детям. Здесь многое зависит и от крепости здоровья матери и от наследуемых ребёнком индивидуальных черт.
Здесь неспецифический вид разделяется и следует по двум путям:
- Абсолютный, с ярко выраженной нечувствительностью к некоторым вирусным бактериям при самых неблагоприятных условиях.
- Относительный, который может быть не так стоек, если создать ему неблагоприятную среду. Примером может служить следующий случай: ребенок сильно переохладился, и находился в данных условиях довольно долго, что повлекло за собой вероятность заболевания ветрянкой. При плюсовой температуре, процент того, что он сможет заболеть, равен нулю.
Есть также понятие личной невосприимчивости к вирусам. Это определяется силой иммунной системы и нормой эндокринных показателей.
Можно наблюдать некоторые расовые особенности и игнорирование инфекций, которые проявляются в виде устойчивости к заболеваниям. Связаны подобные явления и с географическим положением.
Например: жители многих регионов Африки намного устойчивее к такому заболеванию как малярия. Они буквально «на ногах» способны перенести болезнь, тогда как европеец будет лежать с высокой температурой не в силах подняться.
Специфический иммунитет
Что такое специфический вид иммунитета? Он не передается генетическим путем, и формирование его длится всю жизнь. У него также есть свои подгруппы, и условно из них может быть составлена следующая таблица: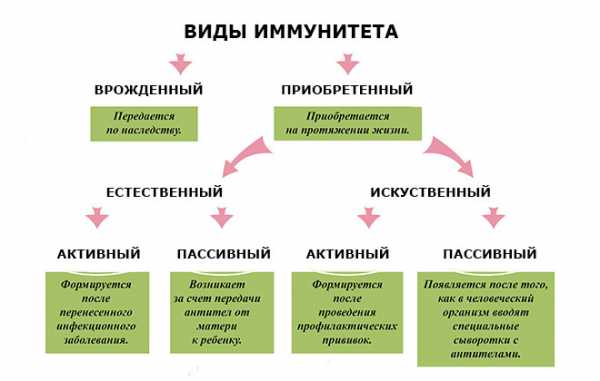
- Специфический или приобретённый естественный активный вид выявляется после того, как человек переболел серьёзной инфекцией. Но не только, в случае нахождения его долгое время в близком контакте с патогенными микробами, также может возникнуть данная форма. Она является достаточно устойчивой. Люди, переболевшие в детстве корью, не смогут заболеть этим недугом ещё раз. По срокам устойчивости данный иммунитет может быть не долговечен, однако сохраняться на десятки лет, постепенно утрачивая свою силу. Можно привести пример с брюшным тифом. При некоторых индивидуальных особенностях, при гриппе, данная форма имеет ещё более кратковременное действие. Если вы переболели этим недугом, то есть большая вероятность, вновь заболеть им через определенный период.
- Специфический, он же приобретенный естественный пассивный вид. Он попадает с передачей иммуноглобулина класса G, от материнской плаценты в организм малыша. Ещё более мощной поддержкой является материнское молоко.
- Специфический искусственный активный. Механизмы появления его в организме человека, связаны с вакцинациями. Устанавливается защита спустя какое – то время после вакцины и может держаться на первых позициях от года до многих лет. Он не имеет возможности передаваться наследственным способом. Особенно востребован при массовых инфекциях губительного свойства.
- Специфический искусственный пассивный. В случае, если иммунитет пассивен к выработке антител, в больной организм вводится уже сформировавшиеся в своей активности белки чужеродного свойства. Зачастую при этой довольно опасной процедуре наблюдается анафилактический шок. По устойчивости этот вид не длителен, до трех недель, затем идет на спад.
Специфические и неспецифические факторы иммунитета имеют отличие и разные способы воздействия на человека. Кроме вышеуказанных видов защиты, наш организм имеет еще массу способов противостояния негативным вторжениям в виде болезнетворных бактерий, микробов и вирусных инфекций.
К барьерным структурам, прежде всего, принадлежат:
- кожная система;
- слизистая оболочка;
- кровяная сыворотка.
Всё продумано до мелочей, любая внешняя атака может быть «отбита» при помощи механических, физико – химических или биологических способов. Остаётся только удивляться и восхищаться, как наш организм способен защищать сам себя.
- Механические проникновения отбиваются кожными покровами и слизистыми оболочками. От укусов насекомых, ожогов и порезов, переломов и травм спасает внешние повреждения и разрыв целостности структур.
- Физико – химические свойства в виде пота и сальных желез приобретают статус защитной реакции, а слизистые выделения, не что иное, как защитное действие на внешние микробные атаки.
- Иммунобиологические свойства начинают свою работу, в случае, если сделан прорыв через кожный слой. Тут же включается защита гуморального и клеточного уровней.
Как не восхищаться продуманностью действий и взаимовыручкой всех внутренних систем и органов друг друга. Этот уникальный мир следует беречь и приумножать его внутренний потенциал. Достаточно получить сбой в одном из звеньев, как может пострадать весь целостный облик.
Берегите своё здоровье и работайте над усилением иммунитета, ведь это ваше главное богатство. Он способно служить вам верой и правдой долгие годы!
Неспецифический, врожденный иммунитет
Элементы врожденного (неспецифического) иммунитета включают анатомические барьеры, секреторные молекулы и клеточные компоненты. Среди механических анатомических барьеров – кожа и эпителиальные слои слизистых, движение кишечника и колебание бронхолегочных ресничек. С этими защитными поверхностями ассоциируются химические и биологические факторы.
Механические факторы как часть неспецифического иммунитета
Эпителиальные поверхности формируют физический барьер, который практически непроницаемый для большинства инфекционных агентов. Таким образом, кожа выступает в качестве нашей первой линией обороны против вторжения патогенов. Десквамации эпителия кожи также помогает удалить бактерии и другие инфекционные агенты, которые присоединяются к эпителиальным поверхностям. Движения ресничек, перистальтика способствуют дыхательным путям и желудочно-кишечному тракту освобождаться от микроорганизмов. Промывающее действие слез и слюны помогает предотвратить инфекции глаз и рта. Захватывающий эффект слизи дыхательных путей и желудочно-кишечного тракта способствует защите легких и пищеварительной системы от инфекции.
Химические факторы в неспецифическом иммунитете
Жирные кислоты пота ингибируют рост бактерий. Лизоцим и фосфолипаза найдены в слезах, слюне и носовых выделениях формируют поры в клеточной стенке бактерий и дестабилизируют мембрану. Низкий рН пота и желудочного секрета предотвращает рост бактерий. Дефензины (низкомолекулярные белки), обнаруженные в легких и желудочно-кишечном тракте, обладают антимикробной активностью. Поверхностно-активные вещества в легких действуют как опсонины (вещества, способствующие фагоцитозу частиц фагоцитами).
Биологические факторы и неспецифический иммунитет
Нормальная флора на коже и в желудочно-кишечном тракте может предотвращать колонизацию патогенных бактерий путем секреции токсичных веществ или конкурируя с патогенными бактериями за питательные вещества или прикрепление к клеточной поверхности.
Гуморальные факторы неспецифического иммунитета
Анатомические барьеры являются очень эффективными в предотвращении колонизации тканей микроорганизмами. Однако при повреждении тканей анатомические барьеры нарушаются, и инфекционные агенты проникают вглубь тканей. В игру включается другой врожденный механизм обороны, а именно гуморальные факторы острой фазы воспаления Гуморальные факторы играют важную роль в организации воспаления. Они находятся в сыворотке или образуются в месте локализации инфекции.
- Система комплемента - Система комплемента является основным гуморальным неспецифическим защитным механизмом. Активация комплемента сопровождается повышением проницаемости сосудов, мобилизацией клеток-фагоцитов, лизисом и опсонизацией бактерий.
- Система свертывания крови - в зависимости от тяжести повреждения тканей, свертывающая система может быть или быть активирована. Некоторые продукты свертывающей системы способствуют неспецифической защите путем повышения сосудистую проницаемость, и действовать в качестве хематрактантов фагоцитирующих клеток. Кроме того, некоторые из продуктов локально активированной системы свертывания обладают антимикробным действием. Например, бета-лизин, белок, продуцируемый тромбоцитами во время свертывания, лизирует многие грамположительные бактерии, действуя как катионный детергент.
- Лактоферрин и трансферрин - связывают железо, необходимое для дыхания бактерий, и таким образом ограничивают рост бактерий.
- Интерфероны - белки, которые могут ограничить репликацию вируса в клетках.
- Лизоцим – фермент, разрушающий клеточную стенку бактерий.
- Интерлейкин-1 (ИЛ-1) – индуцирует лихорадку синтез белков острой фазы, которым присущи плейтропные свойства.
Клеточное звено неспецифического иммунитета
Частью воспалительного ответа на инфекцию является мобилизация полиморфноядерных нейтрофилов, эозинофилов и макрофагов к месту колонии инфекции. Эти клетки являются основной линией обороны в неспецифической иммунной защите.
- Нейтрофилы - полиморфноядерные лейкоциты (ПЯЛ, рис.1), рекрутируются в места инфекции, где они фагоцитируют бактерии и убивают их либо внутриклеточно, либо во внеклеточных ловушках (рис.). Кроме того, ПЯЛ вносят вклад в повреждение тканей, во время воспаления и репарацию их после удаления инфекции.
- Макрофаги - тканевые макрофаги (фигура 5, 6, 7), и моноциты (рис. 4 и 8), которые дифференцируются в макрофаги, выполняют функцию фагоцитоза и внутриклеточное уничтожение микроорганизмов. Кроме того, макрофаги, способны к экстрацеллюлярному киллингу зараженных или поврежденных самих себя. Кроме того, макрофаги принимают участие в ремоделировании тканей. Макрофаги – антиген представляющие клетки: они презентируют процессированный антиген в комплексе с белками ГКГС класса II Т Н1, индукция специфический иммунный ответа.
- Естественные киллеры (ЕК) и лимфокин активированные киллеры (ЛАКК) - ЕК и ЛАКК могут неспецифически убивать инфицированные вирусом и опухолевые клетки. Эти клетки не являются частью воспалительного ответа, но они играют важную роль в неспецифическом иммунитете к вирусным инфекциям и опухолям.
- Эозинофилы - (рис. 6а и б) белки гранул эозинофилов эффективны в убийстве некоторых паразитов.
Неспецифические (естественные) факторы иммунитета
Со времени становления иммунологии как науки возникло представление о врожденном и приобретенном противомикробном иммунитете. Врожденный иммунитет к данной инфекции - это состояние, не зависящее от имевшегося ранее спонтанного или экспериментально вызванного контакта с возбудителем или с его антигенами. Врожденный иммунитет - наследуемая устойчивость животных против определенного возбудителя, в основном породная. Приобретенный иммунитет - невосприимчивость, возникающая у животных в процессе естественного переболевания соответствующей инфекционной болезнью или искусственно созданная у неболевшего (здорового) организма путем его иммунизации биопрепаратами (вакциной, сывороткой).
Таким образом, защита организма от инфекции складывается из последовательного включения в борьбу с проникшим возбудителем трех различных эшелонов этой защиты, составляющих единый функциональный комплекс, включающий в себя:
1) факторы естественной резистентности,
2) ранний индуцибельный ответ,
3) адаптивный или приобретенный иммунный ответ.
Неспецифический (естественный) противомикробный иммунитет обеспечивают следующие факторы:
1) анатомофизиологические;
2) гуморальные;
3) клеточные.
Анатомофизиологические факторы неспецифической резистентности
Микроорганизмом в ходе эволюции были выработаны защитные механизмы от возбудителя. Для проникновения в восприимчивые клетки и ткани микробу необходимо преодолеть немало защитных барьеров.
Кожно-слизистые барьеры. Первую атаку микроба-агрессора испытывают неповрежденные кожа и слизистые оболочки, это не только механическая преграда, но и стерилизующий фактор в отношении многих видов микроорганизмов. Кожа и слизистые оболочки покрыты слоем эпителиальных постоянно обновляющихся клеток, преграждающих путь микробу. Вместе с ними при постоянно происходящем слущивании эпителия удаляется и осевшая на нем патогенная микрофлора. На чистой коже относительно быстро погибают некоторые виды бактерий. Например, культура чудесной палочки, нанесенная на здоровую кожу человека, погибает спустя 20 мин. Бактерицидное действие кожи обусловлено веществами, выделяемыми потовыми и сальными железами, а также содержащимися в коже жирными кислотами.
Выраженными барьерными свойствами обладают также конъюнктива, слизистая оболочка носа и др.
Секреты желез пищеварительного тракта наряду со своими физиологическими функциями обладают способностью обезвреживать многих болезнетворных микробов. Слюна - первый секрет, воздействующий на пищевые вещества, а также микрофлору, поступающие в ротовую полость. Желудочный сок также обладает бактерицидным и бактериостатическим действием на ряд видов патогенных микробов, однако в содержимом желудка выживают туберкулезные и спорообразующие бактерии. Желчь действует бактериостатически на ряд микроорганизмов. Слезная жидкость оказывает бактерицидное действие на патогенные микробы, особенно из группы кокков.
Болезнетворные микробы, преодолевшие кожный и слизистый барьеры, начинают массированное проникновение в ткани. В инфицированном участке огромная масса фагоцитов создает защитный вал вокруг микроорганизмов. Возникает местный воспалительный процесс, который ограничивает распространение микробов в соседние ткани и кровь. И. И. Мечников первым указал на защитную роль воспаления.
Воспаление - сложная сосудисто-тканевая защитно-приспособительная реакция организма на действие патогенного раздражителя. Воспаление выполняет защиту организма от воздействия патогенного фактора в виде специфических реакций - фагоцитоза и выработки антител. Благодаря воспалительной реакции фокус повреждения отграничивается от всего организма, происходит ликвидация патогенного фактора, повышение местного и общего иммунитета. При определенных условиях воспаление может приобретать иногда вредоносное значение для организма (некроз тканей, нарушение функций).
Лимфатическая система. При дальнейшем продвижении в ткани и кровь микробы встречают новый барьер - лимфатические узлы. Они расположены по ходу лимфатических сосудов и играют роль своеобразных фильтров, которые задерживают микробные клетки и другие нерастворимые частицы.
Если возбудителю удается преодолеть и эти защитные преграды, то в микроорганизме происходит изменение уровня обмена веществ и определенных физиологических процессов. Так, при многих инфекциях повышается температура тела в связи с усилением обменных и энергетических процессов. Лихорадочную реакцию, особенно в начальной фазе болезни, следует рассматривать кик защитную. Установлено губительное действие повышенной температуры (38...40°С) на некоторые вирусы. Повышение мочеотделения и потоотделения способствуют освобождению организма от ряда возбудителей. При кишечных болезнях диарея сопровождается обильным выделением из организма возбудителей.
14.Видовой (наследственный) иммунитет. Неспецифические факторы защиты.
Врожденный, иди видовой, иммунитет, он же наследственный, генетический, конституциональный — это выработанная в процессе филогенеза генетически закрепленная, передающаяся по наследству невосприимчивость данного вида и его индивидов к какому-либо антигену (или микроорганизму), обусловленная биологическими особенностями самого организма, свойствами данного антигена, а также особенностями их взаимодействия.
Примером может служить невосприимчивость человека к некоторым возбудителям, в том числе к особо опасным для сельскохозяйственных животных (чума крупного рогатого скота, болезнь Ньюкасла, поражающая птиц, оспа лошадей и др.), нечувствительность человека к бактериофагам, поражающим клетки бактерий. К генетическому иммунитету можно также отнести отсутствие взаимных иммунных реакций на тканевые антигены у однояйцовых близнецов; различают чувствительность к одним и тем же антигенам у различных линий животных, т. е. животных с различным генотипом.
Объяснить видовой иммунитет можно с разных позиций, прежде всего отсутствием у того или иного вида рецепторного аппарата, обеспечивающего первый этап взаимодействия данного антигена с клетками или молекулами-мишенями, определяющими запуск патологического процесса или активацию иммунной системы. Не исключены также возможность быстрой деструкции антигена, например, ферментами организма или же отсутствие условий для приживления и размножения микроба (бактерий, вирусов) в организме. В конечном итоге это обусловлено генетическими особенностями вида, в частности отсутствием генов иммунного ответа к данному антигену.
Видовой иммунитет может быть абсолютным и относительным. Например, нечувствительные к столбнячному токсину лягушки могут реагировать на его введение, если повысить температуру их тела. Белые мыши, не чувствительные к какому-либо антигену, приобретают способность реагировать на него, если воздействовать на них иммунодепрессантами или удалить у них центральный орган иммунитета — тимус.
Неспецифические факторы защиты
Противоинфекционную защиту осуществляют:
1) кожа и слизистые оболочки;
2) лимфатические узлы;
3) лизоцим и другие ферменты полости рта и ЖКТ;
4) нормальная микрофлора;
5) воспаление;
6) фагоцитирующие клетки;
7) естественные киллеры;
8) система комплемента;
9) интерфероны.
Неповрежденная кожа и слизистые оболочки являются барьером, препятствующим проникновению микроорганизмов внутрь организма. В результате слущивания эпидермиса удаляются многие транзиторные микроорганизмы. Бактерицидными свойствами обладает секрет потовых и сальных желез. При наличии травм, ожогов кожа формирует входные ворота для инфекции.
Секреты, выделяемые слизистыми оболочками, слюнными и пищеварительными железами, слезы смывают микроорганизмы с поверхности слизистых, оказывают бактерицидное действие.
Лизоцим – белок, содержащийся в тканевых жидкостях, плазме, сыворотке крови, лейкоцитах, материнском молоке и др. Он вызывает лизис бактерий, неактивен в отношении вирусов.
Представители нормальной микрофлоры могут выступать в качестве антагонистов патогенных микроорганизмов, препятствуя их внедрению и размножению.
Неспецифические факторы иммунитета микробиология. Неспецифические факторы иммунитета
Различают видовой и приобретенный иммунитет.
Видовой иммунитет передается но наследству, характерен для данного вида. Например, человек невосприимчив к чуме рогатого скота, к куриной холере, собаки невосприимчивы к туберкулезу. Видовой иммунитет неспецифичен, то есть одни и те же защитные механизмы действуют против разных видов микробов. Это наиболее прочный вид иммунитета.
Неспецифические факторы естественной резистентности защищают организм от микробов при первой встрече с ними. Эти же факторы участвуют и в формировании приобретенного иммунитета.
Ареактивность клеток является наиболее стойким фактором естественной защиты. При отсутствии клеток, чувствительных к данному микробу, токсину, вирусу организм полностью защищен от них. Так, например, крысы нечувствительны к дифтерийному токсину.
Кожа и слизистые оболочки представляют собой механический барьер для большинства патогенных микробов. Кроме того, на микробы губительно действуют выделения потовых и сальных желез, содержащие молочную и жирные кислоты. Чистая кожа обладает более сильными бактерицидными свойствами. Удалению микробов с кожи способствует слущивание эпителия.
В секретах слизистых оболочек содержится лизоцим (lysozyme) -фермент, лизирующий клеточную стенку бактерий, главным образом, грамположительных. Лизоцим содержится в слюне, секрете конъюнктивы, а также в крови, в макрофагах, в слизи кишечника. Открыт впервые П.Н. Лащенковым в 1909 г. в белке куриного яйца.
Эпителий слизистых оболочек дыхательных путей является препятствием для проникновения патогенных микробов в организм. Частицы пыли и капли жидкости выбрасываются наружу со слизью, выделяющейся из носа. Из бронхов и трахеи попавшие сюда частицы выводятся движением ресничек эпителия, направленным кнаружи. Эта функция мерцательного эпителия обычно нарушена у злостных курильщиков. Немногие частички пыли и микробы, достигшие легочных альвеол, захватываются фагоцитами и обезвреживаются.
Секрет пищеварительных желез. Желудочный сок губительно действует на микробов, поступающих с водой и пищей, благодаря наличию соляной кислоты и ферментов. Пониженная кислотность желудочного сока способствует ослаблению сопротивляемости к кишечным инфекциям, таким как холера, брюшной тиф, дизентерия. Бактерицидным действием обладают также желчь и ферменты кишечного содержимого.
Лимфатические узлы. Микробы, проникшие через кожу и слизистые оболочки, задерживаются в регионарных лимфатических узлах. Здесь они подвергаются фагоцитозу. В лимфатических узлах также содержатся так называемые нормальные (естественные) киллеры-лимфоциты (англ, killer - убийца), несущие функцию противоопухолевого надзора - разрушение собственных клеток организма, измененных вследствие мутаций, а также клеток, содержащих вирусы. В отличие от иммуных лимфоцитов, формирующихся в результате иммунного ответа, естественные киллеры распознают чужеродные агенты без предварительного контакта с ними.
Воспаление (сосудисто-клеточная реакция)- одна из филогенетически древних защитных реакций. В ответ на проникновение микробов формируется местный воспалительный очаг в результате сложных изменений микроциркуляции, системы крови и клеток соединительной ткани. Воспалительная реакция способствует удалению микробов или задерживает их развитие и поэтому играет защитную роль. Но в ряде случаев, при повторном попадании агента, вызвавшего воспаление, оно может принять характер повреждающей реакции.
Фагоцитоз - процесс активного поглощения клетками организма микробов и других чужеродных частиц (греч. phagos - пожирающий + kytos - клетка), в том числе собственных погибших клеток организма. И.И. Мечников - автор фагоцитарной теории иммунитета - показал, что явление фагоцитоза - это проявление внутриклеточного переваривания, которое у низших животных, например, у амеб, является способом питания, а у высших организмов фагоцитоз является механизмом защиты. Фагоциты освобождают организм от микробов, а также уничтожают старые клетки собственного организма.
По Мечникову, все фагоцитирующие клетки подразделяются на макрофаги и микрофаги. К микрофагам относятся полиморфноядерные гранулоциты крови: нейтрофилы, базофилы, эозинофилы. Макрофаги - это моноциты крови (свободные макрофаги) и макрофаги различных тканей организма (фиксированны
5. Гуморальные механизмы неспецифического иммунитета
Гуморальные механизмы неспецифической иммунной защитыобеспечиваютсяспециальными гуморальными факторами(преимущественнобелковой природы), которые
находятся в плазме крови(т.е. постоянно предсуществуют в крови независимо от проникновения чужеродных агентов в организм),
могут продуцироваться поврежденными клетками тканей организмаиэндотелием сосудистой стенки,
либо вырабатываются активированными фагоцитами и другими разновидностями лейкоцитов.
Эти факторы неспецифической гуморальной защиты организма от генетически чужеродных структур способны оказывать
опсонизирующее действиев отношении бактерий и вирусов (т.е. облегчают фагоцитоз чужеродных структур фагоцитами),
батериолитическое ивиролитическое действие,
некоторые из этих белков являются хемоаттрактантами(вызывают хемотаксис лейкоцитов в очаг воспаления) имедиаторами воспаления.
К гуморальным факторам, обеспечивающим неспецифическую иммунную защиту, относят:
систему комплемента,
т.н. белки острой фазы,
медиаторы воспаления, некоторые из которых образуется в результате активации системы комплемента, другие – продуцируются поврежденными собственными клетками организма, третьи – являются продуктами жизнедеятельности или составными компонентами патогенных организмов.
5.1. Краткая характеристика системы комплемента
Система комплементапредставляет собой большую группу белков плазмы крови (около 20 белков)
часть из которых (9 белков, обозначаемых буквой Си имеющих определенный порядковый номер соответственно последовательности их открытия (но не активации): С1, С2, С3 и т.д. вплоть до С9) являютсянеактивными ферментамии активируются в определенной последовательности путемограниченного протеолиза,
тогда как другие белки системы комплемента выступают в роли регуляторных факторов.
В результате последовательной активации компонентов системы комплементаобразуются:
медиаторы воспаления,
факторы, оказывающие опсонизирующее действие в отношении бактерий и вирусов,
литические комплексы, способные убивать антигены, связанные с антителами.
Начало познанию механизмов функционирования системы комплемента было положено французом Жюлем Борде, работавшим в лаборатории И. Мечникова в Париже, в конце XIXвека. Окончательное название этой системы (комплемент) дано Паулем Эрлихом. Некоторые из белков системы комплемента синтезируются гепатоцитами печени, другие – клетками эпителия кишечника и макрофагами. Большинство факторов системы комплемента представляют собойнеактивные ферменты(проферменты или зимогены), находящиеся в плазме крови в неактивной форме и активирующиеся в результате ограниченного протеолиза (отщепления от них ингибиторного фрагмента). Причем активация компонентов системы комплемента осуществляются по типу цепного каскадного ферментативного процесса, при котором один активировавшийся фактор этой системы (активный фермент) путем ограниченного протеолиза другого фактора приводит к его активации, в результате которой этот другой фактор превращается в активный фермент и катализирует активацию следующего компонента системы комплемента. Каскадный механизм активации системы комплемента обеспечивает формированиебыстрого многократно усиленного ответа на первичный сигнал, поскольку каждый фактор этой системы, становясь активным, катализирует активацию большого количества молекул другого фактора, а те, в свою очередь, активируют еще большее количество молекул следующего фактора и т.д.
В результате ограниченного протеолизавсех компонентов системы комплемента образуетсяпо 2 фрагмента каждого компонента(т.е. каждый компонент расщепляется на две части), один из которых являетсяболее мелким и обозначается буквойа, а другой –более крупным(обозначается буквойb). Например, при активации фактора С3, предполагающей его ограниченный протеолиз, образуется два его фрагмента: С3а (более короткий и низкомолекулярный) и С3b(более длинный и высокомолекулярный).
Например,
ограниченный протеолиз под действием активированного фактора С4b
С2а
С2
С2b
в комплексе с С4b и ионами Мg2+
ограниченный протеолиз
С3
С3а
С3b
Мелкие фрагментыкомпонентов комплемента (обозначаемые буквойа), как правило, ферментативной активностью не обладают и поэтому не принимают участия в активации следующего компонента системы комплемента, но зато эти фрагменты
являются хемоаттрактантами(т.е. оказывают хемотаксическое действие в отношении лейкоцитов),
способны повышать проницаемость сосудистой стенкииплазматических мембран клеток,
кроме того, они активируют нейтрофилы и макрофаги
являются медиаторами воспаления(т.е. вызывают некоторые проявления воспалительных реакций).
Крупные фрагментыкомпонентов системы комплемента проявляют ферментативную активность. Причем в большинстве своем они являютсясериновыми протеазамии гидролизуют пептидные связи между сериновыми аминокислотными остатками в молекулах других компонентов системы комплемента. В результате такого гидролиза компонент комплемента расщепляется на два фрагмента: один из которых, более мелкий, не проявляет ферментативной активности, а другой – является активной протеазой, расщепляющей следующий компонент и таким образом активирующий его.
Результатом каскадного механизма последовательной активации компонентов системы комплементаявляется образованиебелковых комплексов С4b-С3b(связывается с мембранами чужеродных клеток и облегчает фагоцитоз этих клеток нейтрофилами и макрофагами), а такжекомплекса С5b-С9(образует поры в мембранах чужеродных клеток, ассоциированных с антителами, вызывает повреждение и уничтожение этих клеток).
Различают два пути активации системы комплемента:
классический путь(запускается под влиянием иммунных комплексов – комплексов "антиген-антитело")
альтернативный путь(запускается только лишь каким-то патогеном без участия антител, происходит гораздо быстрее классического пути активации, поскольку не зависит от первоначальной продукции антител и связывания ими антигенов, а запускается сразу после проникновения патогена в организм).
Эти два пути активации системы комплемента отличаются не только характером запускающих их механизмов и быстротой реакции, но и начальными стадиями этого процесса. Вместе с тем, заключительные этапы и результат активации системы комплемента являются общими для классического и альтернативного путей.
Классический путь активации комплементаначинается с активациииммунными комплексами(комплексами "антиген-антитело")фактора С1. Этот фактор состоит из трех белков:С1q, C1r, C1s.Белок С1q, в свою очередь, состоит из6 субъединиц, в каждой из которых имеетсяглобулярная часть(напоминающая булаву) инитчатая часть. Нитчатые части этих 6 субъединиц в составе молекулы С1qрасположены таким образом, что формируют единую фибриллярную рукоять (коллагеноподобную рукоять), от которой радиально отходит 6 глобулярных булав. Нитчатые компоненты субъединиц белка С1qближе к своему основаниюв присутствии ионов кальциявзаимодействуют с белкамиC1rиC1s(имеют глобулярную форму), так что формируется единый компонент комплемента С1. Глобулярные части белка С1qобладают способностью связываться с иммуноглобулинами класса М иG(сFс-фрагментами иммуноглобулинов, которые являются частями константных областей антител и способны специфически связываться с рецепторами дляFс-фрагментов, расположенными на поверхности некоторых клеток организма – гранулоцитов, лимфоцитов, макрофагов и некоторых других). Взаимодействие белка С1qс антителом происходит только после прикрепления этого антитела к антигену (в связи с тем, что под действием присоединившегося к антителу антигена изменятся пространственная структура антитела, и оно приобретает способность взаимодействовать с белком С1qкомпонента С1). Белок С1qсо свободными антителами не взаимодействует. Причем, если белок С1qможет прикрепляться к единичным молекулам иммуноглобулинов М, связанных с определенными антигенами, то для взаимодействия с антителами классаGнеобходимо несколько молекул этих антител, адсорбированных на поверхности корпускулярного антигена (контакт белка С1qтолько с одной молекулой иммуноглобулинаGне обеспечивает активации компонента С1).
В результате взаимодействия ассоциированных с антигенами антител класса М и Gс белковым компонентом С1qпроисходит некоторое изменение его пространственной структуры, сопровождающееся появлением у него ферментативной активности. Активированный белок С1qмодифицирует белок С1s, придавая ему свойства сериновой протеазы. С приобретением ферментативных свойств белком С1sзавершается первый этап классического пути активации системы комплемента.
Следующий этап предполагает активацию фактора С4 под действием образовавшейся на первом этапе сериновой протеазы (активного белка-фермента С1s). Причем С4 под действием активного белка С1sрасщепляется на два фрагмента: С4а (мелкий, является медиатором воспаления) и С4b(более крупный, ковалентно связывается с мембраной патогена, после чего взаимодействует с фактором С2). После взаимодействия С4bс фактором С2, которое происходит с участием ионов Мg2+, изменятся пространственная структура фактора С2, в результате чего он становится чувствительным к сериновой протеазе (активированному фактору С1s). Под действием этой протеазы фактор С2 расщепляется на два фрагмента: С2а и С2b. Фрагмент С2b, подобно белку С1s, проявляет свойства сериновой протеазы и совместно с белком С4bпредставляет собой фермент С3/С5-конвертазу, прикрепленную к поверхности патогена. Образовавшийся фермент С3/С5-конвертаза взаимодействует с фактором С3 и катализирует расщепление этого фактора на два фрагмента С3а (более мелкий фрагмент, является медиатором воспалительной реакции) и С3b(остается прикрепленным к поверхности патогена). Фрагмент С3bвзаимодействует с определенными рецепторами на фагоцитах, тем самым способствуя прикреплению патогена, с которым он связан, к мембране фагоцита (т.е. фагоцит соединяется с патогенном через посредство компонента С3b). Такое прикрепление патогена к поверхности фагоцита, осуществляющееся с участием компонента С3b, в свою очередь, сопровождается фагоцитозом этого патогена (т.е. компонент С3bоказываетопсонизирующее действиев отношении многих бактерий и вирусов, облегчая их фагоцитоз нейтрофилами и макрофагами). Кроме опсонизирующего действия, компонент С3b, фиксированный на мембране патогена, взаимодействует с фактором С5, делая его доступным для действия С3/С5-конвертазы, которая расщепляет С5 на два фрагмента: С5а (выступает в качестве воспалительного фактора и хемоаттрактанта, привлекая в зону проникновения патогена фагоцитирующие клетки) и С5b(остается связанным с мембраной патогена). Компонент С5bпоследовательно взаимодействует с факторами С6, С7 и С8. Образующийся комплекс С5b678, зафиксированный на мембране патогена, взаимодействует с несколькими молекулами (2 или более) фактора С9, которые под воздействием этого комплекса изменяет свою пространственную структуру так, что как бы развертываются и становятся способными проникать внутрь билипидного слоя мембраны патогена. Внутри билипидного слоя мембраны патогена проникшие молекулы фактора С9 полимеризуются в кольцеобразный мембраноатакующий комплекс, который формирует трансмембранный канал полностью проницаемый для электролитов и воды. Таким образом, комплекс С5b678 способствует проникновению в толщу мембраны патогена молекул фактора С9, которые соединяясь друг с другом, образуют поры в мембране. Количество таких пор в мембране патогена будет тем большим, чем больше проникло молекул фактора С9 в толщу билипидного слоя. За счет высокого коллоидно-осмотического давления внутри чужеродной клетки в нее через поры, образовавшиеся из молекул фактора С9, поступают ионы натрия и вода, что приводит к набуханию клетки и последующему ее лизису.
Таким образом, активация факторов системы комплемента по классическом пути осуществляется в следующей последовательности:
С1 С4 С2 С3 С5 С6 С7 С8 С9
СХЕМА ВЗАИМОДЕЙСТВИЯ И АКТИВАЦИИ КОМПОНЕНТОВ СИСТЕМЫ КОМПЛЕМЕНТА ПО КЛАССИЧЕСКОМУ ПУТИ
С1qrs
Лекция № 10. Иммунитет, виды и формы. Структура иммунной системы. Факторы неспецифической защиты.
Первоначально иммунология возникла как наука о невосприимчивости (иммунитете) к инфекционным болезням. Наиболее существенный вклад в ее создание внесли И.И.Мечников (фагоцитарная или клеточная теория иммунитета) и П.Эрлих (гуморальная теория), в творческой дискуссии между которыми совершенствовались представления об иммунитете.
В настоящее время считается, что наследственный (врожденный, видовой) и приобретенный иммунитет зависит от согласованной деятельности пяти основных систем : макрофагов, комплемента, интерферонов, Т- и В- лимфоцитов, главной системы гистосовместимости (МНС- в английском варианте), обеспечивающих различные формы иммунного ответа.
В современном понимании иммунология- это не только наука, изучающая защиту от инфекционных заболеваний. Иммунология- наука, изучающая механизмы самозащиты организма от всего генетически чужеродного, поддержания структурной и функциональной целостности организма (гомеостаза организма). Подробнее - см. лекцию 1.
Центральным биологическим механизмом иммунитета является механизм распознавания “своего” и “чужого”. Пример- необходимость защиты от собственных мутантных и раковых клеток (одномоментно в организме находится около 10 млн. измененных клеток).
Иммунитет- целостная система биологических механизмов самозащиты организма, с помощью которых он распознает и уничтожает все чужеродное (генетически отличающееся).
Выделяют две основные формы иммунитета- видовой (врожденный) и приобретенный. Приобретенный иммунитет может быть естественный (результат встречи с возбудителем) и искусственный (иммунизация), активный (вырабатываемый) и пассивный (получаемый), стерильный (без наличия возбудителя) и нестерильный (существующий в присутствии возбудителя в организме), гуморальный и клеточный, системный и местный, по направленности- антибактериальный, антивирусный, антитоксический, противоопухолевый, антитрансплантационный.
В основе видового иммунитета лежат различные механизмы естественной неспецифической резистентности. Среди них- кожные покровы и слизистые оболочки, нормальная микрофлора организма, фагоцитоз, воспаление, лихорадка, система комплемента, барьерные механизмы лимфоузлов, противомикробные вещества, выделительные системы организма, главная система гистосовместимости.
Кожа и слизистые- первая линия защиты против возбудителей. Кроме функции механического (анатомического) барьера кожа обладает бактерицидной активностью. Слизь, лизоцим, желудочный сок, слезная жидкость, слюна, деятельность мерцательного эпителия способствует защите слизистых оболочек.
Нормальная микрофлора организма препятствует колонизации организма посторонней микрофлорой (конкуренция за субстраты, различные формы антагонизма, в т.ч. выделение антибиотических веществ, изменение рН и др.).
Фагоцитоз и система комплемента- вторая линия защиты организма против микроорганизмов, преодолевших поверхностные барьеры. Клеточные факторы системы видовой резистентности- фагоциты, поглощающие и разрушающие патогенные микроорганизмы и другой генетически чужеродный материал. Представлены полиморфоядерными лейкоцитами или гранулоцитами- нейтрофилами, эозинофилами и базофилами (клетками миелопоэтического ряда), а также моноцитами и тканевыми макрофагами (клетками макрофагально- моноцитарной системы).
Значение фагоцитирующих клеток для защиты организма впервые доказал И.И.Мечников, разработавший фагоцитарную теорию иммунитета.
Стадии фагоцитоза.
Процесс фагоцитоза (поглощения твердофазного объекта) состоит из пяти стадий.
1.Активация (усиление энергетического метаболизма). Факторами активации и хемотаксиса являются бактериальные продукды (ЛПС, пептиды), компоненты комплемента (С3 и С5), цитокины и антитела.
2.Хемотаксис.
3.Адгезия.
4.Поглощение.
5.Исход фагоцитоза.
Адгезия связана с наличием ряда рецепторов на поверхности фагоцитов ( к Fc- фрагментам антител, компонентам комплемента, фибронектину), обеспечивающих прочность рецептор- опосредованных взаимодействий опсонинов, обволакивающих микроорганизмы и ограничивающих их подвижность (антитела, С3в, фибронектин).
Фагоциты обладают амебоподобными псевдоподиями. При поглощении образуется фагосома с поглощенным объектом (бактерией), к ней присоединяется и сливается содержащая литические ферменты лизосома, образуется фаголизосома.
Возможно три исхода фагоцитоза:
- завершенный фагоцитоз;
- незавершенный фагоцитоз;
- процессинг антигенов.
Завершенный фагоцитоз- полное переваривание микроорганизмов в клетке- фагоците.
Незавершенный фагоцитоз- выживание и даже размножение микроорганизмов в фагоците. Это характерно для факультативных и особенно - облигатных внутриклеточных паразитов. Механизмы персистирования в фагоцитах связаны с блокадой фагосомо- лизосомального слияния (вирус гриппа, микобактерии, токсоплазмы), резистентностью к действию лизосомальных ферментов (гонококки, стафилококки), способностью микробов быстро покидать фагосомы после поглощения и длительно пребывать в цитоплазме (риккетсии).
В процессе фагоцитоза происходит “окислительный взрыв” с образованием активных форм кислорода, что обеспечивает бактерицидный эффект.
К одной из важнейших функций макрофагов (наряду с хемотаксисом, фагоцитозом, секрецией биологически активных веществ) является переработка (процессинг) антигена и представление его иммунокомпетентным клеткам с участием белков главной системы гистосовместимости (МНС) класса 2.
Фагоцитоз- не только уничтожение чужеродного, но и представление антигена для запуска иммунных реакций и секреции медиаторов иммунных и воспалительных реакций. Система макрофагов- центральное звено не только естественной резистентности (видового иммунитета), но и играет важную роль в приобретенном иммунитете, кооперации клеток в иммунном ответе.
Воспаление как защитная реакция организма на различные повреждения тканей возникло на более высокой ступени эволюции, чем фагоцитоз и характерно для высокоорганизованных организмов, обладающих кровеносной и нервной системами.
Инфекционное воспаление сопровождается различными сосудистыми и клеточными (включая фагоцитоз) реакциями, а также запуском целого ряда медиаторов воспалительных реакций (гистамина, серотонина, кининов, белков острой фазы воспалеия, лейкотриенов и простагландинов, цитокинов, системы комплемента).
Многие бактериальные продукты активируют клетки макрофагально- моноцитарной системы и лимфоциты, отвечающие на них выделением биологически активных продуктов- цитокинов, в частности интерлейкинов. Их можно характеризовать как медиаторы клеточных иммунных реакций. В воспалительных реакциях основную роль имеет интерлейкин-1 (ИЛ-1), стимулирующий лихорадку, повышающий проницаемость сосудов и адгезивные свойства эндотелия, активирующий фагоциты.
Лихорадка. Повышение температуры тела- защитная реакция организма, ухудшающая условия для размножения многих микроорганизмов, активирует макрофаги, ускоряет кровоток и усиливает обменные процессы в организме.
Барьерные функции лимфоузлов. По выражению П.Ф.Здродовского (1969) лимфоузлы- своеобразный биологический фильтр для возбудителей, переносимых с лимфой. Здесь проникшие через кожу или слизистые и занесенные током лимфы микроорганизмы задерживаются и подвергаются действию макрофагов и активированных лимфоцитов.
Система комплемента- комплекс белков и гликопротеидов сыворотки крови человека и позвоночных животных (их более 20). Отдельные компоненты опосредуют процессы воспаления, опсонизацию чужеродных фрагментов для последующего фагоцитоза, участвуют наряду с макрофагами в непосредственном уничтожении микроорганизмов и других чужеродных клеток (лизис бактерий и вирусов). В условиях физиологической нормы компоненты системы комплемента находятся в неактивной форме. Известны три пути активации системы комплемента- классический, альтернативный и с использованием С1- шунта.
Классический путь- каскад протеазных реакций с компонента С1q до С9, реализуется при наличии антител к соответствующему антигену. С комплексом “антиген- антитела” взаимодействует компонент С1q, затем С4, следом- С2. Образуется комплекс “антиген- антитела-С1С4С2”, с ним соединяется С3 (центральный компонент системы) и запускается цепь активации с эффекторными функциями (опсонизация и лизис бактерий, активация системы макрофагов, воспаление).
Альтернативный путь реализуется при первичном контакте с возбудителем (когда еще нет антител). Он индуцируется ЛПС и другими микробными антигенами. С1, С4, С2 не участвуют, альтернативный и классический пути смыкаются на уровне С3.
Гуморальные факторы специфического иммунитета
этого белка основано на конкуренции с микроорганизмами за присоединение железа. Известно, что при избытке железа вирулентность некоторых видов микроорганизмов (стрептококк и кандида) резко возрастает. Происхождение лактоферина в полости рта мало изучено.
Большое значение в формировании неспецифичной противоинфекционной резистентности слизистой оболочки полости рта, противовирусной, принадлежит интерферону. Следует отметить, что интерферон может уг. нетать реакции гиперчувствительности замедленного типа. Синтезируется интерферон лимфоцитами, макрофагами и фибробластами. При вирусной инфекции клетки синтезируют интерферон и секретируют его в межклеточное пространство, где он связывается со специфическими рецепторами соседних непораженных клеток.
Результат действия интерферона состоит в образовании барьера из неинфицированных клеток вокруг очага вирусной инфекции с целью ограничения ее распространения. Интерфероны играют важную роль в борьбе с вирусами, а не в предотвращении вирусной инфекции. В последнее время получены данные, свидетельствующие о том, что интерфероны. как антагонисты онкобелка, ингибируют пролиферативную активность клеток.
К числу факторов неспецифической защиты слизистой оболочки полости рта можно отнести комплемент (С) — сложный комплекс белков. Комплемент в полости рта в основном находится в зубодесневой жидкости и обуславливает острую воспалительную реакцию тканей десны, уничтожение микробов и повреждение тканей.
Кроме общих факторов неспецифической защиты, важную протектив-ную роль играют ферменты слюны, такие как амилаза, щелочная и кислая фосфатаза, РНК-аза, ДНК-аза, протеолитические ферменты и ингибиторы протеолиза. К активу защитных факторов полости рта имеет смысл отнести эндогенные пирогены, которые выделяются фагоцитирующими макрофагами при вирусных заболеваниях, а также систему пропердина.
Таким образом, слюна представлена практически полным набором ферментов, способных разрушать практически все виды простых биологических субстратов (белки, жиры, углеводы).
Клеточные факторы неспецифической резистентности
В полости рта клеточные реакции неспецифической защиты осуществляются преимущественно полинуклеарными нейтрофилами и макрофагами. Макрофаги представлены в собственном слое слизистой оболочки гистиоцитами, в то время как нейтрофилы в большом количестве выявляются в слюне и зубодесневой борозде.
Гистиоциты (оседлые макрофаги) в отличие от микрофагов являются длительно существующими клетками, функция которых сводится к борьбе теми бактериями, вирусами и простейшими, которые способны существовать внутри клетки хозяина. Макрофаги, которые пассивны в слизистой оболочке полости рта, активизируются в процессе развития воспаления.
у больных кариесом зубов и пародонтитом выявлены разнообразные изменения неспецифических факторов местного и системного иммунитета.
Данные о содержании лизоцима в сыворотке крови и слюне больных кариесом разнообразны. По данным большинства исследователей, содержание и активность лизоцима сыворотки крови при кариесе зубов явно уменьшается, причем у лиц с острейшим течением заболевания активность этого фермента снижается значительно. Данные других авторов не подтверждают существование зависимости возникновения кариеса зубов от содержания лизоцима в крови. Содержание лизоцима в слюне, по данным ряда исследователей, снижается по мере усиления активности кариозного процесса, активность лизоцима в смешанной слюне достоверно снижена при остром кариесе. Другими исследователями выявлена противоположная тенденция: увеличение титра лизоцима в слюне при неослож-ненном кариесе.
При пародонтите уровень лизоцима как в слюне, так и в жидкости зу-бодесневого кармана больных снижается уже на начальных стадиях заболевания. У больных с выраженным эксудативным процессом в тканях пародонта выявлена высокая протеолитическая активность слюны и десневой жидкости.
Таким образом, при кариесе зубов и пародонтите имеет место несостоятельность многих факторов неспецифической антиинфекционной резистентности, особенно местной, в полости рта.
Формирование гуморальной специфической защитной реакции на антиген обеспечивает В-звено иммунной системы.
Основным гуморальным фактором местной антиинфекционной резистентности полости рта являются IgA-антитела, в частности секреторные. Источники IgA-слюны — малые и большие слюнные железы. Считается, что их основное защитное свойство обусловлено способностью непосредственно действовать на бактерии, вызывая их агглютинацию и мобилизацию, Ig-A слюны препятствуют адгезии микроорганизмов, в том числе грибков и вирусов к поверхности слизистой оболочки полости рта, а также к твердым тканям зуба. Кроме этого, они могут ограничивать образование колоний и снижать вирулентность возбудителей инфекции.
56
57
Иммуноглобулин А также имеет большое значение в регуляции микрофлоры в полости рта. ее расселении и поступлении внутрь тканей. Недостаток его в слюне может привести к нарушениям соотношения между микрофлорой полости рта. особенно ее условно патогенными формами и микроорганизмами.
Нарушение барьерной функции IgA-секретов может быть причиной многих аллергических заболеваний, развития клеточных иммунных реакций с повреждением слизистых оболочек.
Клеточные факторы специфического иммунитета
Клеточно-опосредованные реакции иммунитета осуществляются T-лимфоцитами, популяция их неоднородна и представлена специализированными по функциям клетками.
На поверхности слизистой оболочки полости рта Т-лимфоциты встречаются лишь в жидкости десневой борозды. На других участках свою функцию они осуществляют в собственной пластинке слизистой оболочки.
Следует отметить, что в полости рта ткани десны наиболее насыщены Т-лимфоцитами. Они продуцируют фактор, стимулирующий функцию остеокластов, которые усиливают резорбцию костной ткани альвеолярного отростка.
Функциональная анатомия височно-нижнечелюстного сустава в возрастном аспекте
Нормальная функция височно-нижнечелюстного сустава (ВНЧС) зависит от правильных взаимоотношений суставных поверхностей костей, эластичности тканей, образующих сустав, расположения и состояния внутрисуставного диска, состояния хрящей, покрывающих суставные поверхности, функционального состояния синовиального слоя капсулы и состава синовиальной жидкости, а также слаженности работы нервно-мышечного аппарата. Поэтому знание анатомических особенностей и биомеханики ВНЧС необходимо для правильного понимания патогенеза различных заболеваний, их профилактики, четкой диагностики, рационального подхода к лечению.
ВНЧС имеет много общего с другими синовиальными суставами, однако ряд следующих анатомических и функциональных характеристик отличают его от других суставов:
а) суставные поверхности костей покрыты фиброзной тканью — волокнистым хрящом, а не гиалиновым;
б) нижняя челюсть содержит зубы, их форма и расположение в кости влияют на характер движения суставов;
в) левый и правый суставы функционируют вместе как единое целое, и любое движение в одном их них отражается на характере движения в другом;
г) полная зависимость внутрисуставных взаимоотношении от харак- тера смыкания зубных рядов (окклюзии) и состояния жевательных мышц;
д) суставная капсула прикрепляется внутри нижнечелюстной ямки, а не вне суставной ямки, как в других суставах;
ж) наличие внутрисуставного диска. Элементы ВНЧС (рис. 25):
головка нижней челюсти;
нижнечелюстная ямка височной кости;
суставной бугорок височной кости;
позадисуставной конус;
внутрисуставной диск;
капсула сустава;
внутри- и внесуставные связки;
синовиальная жидкость.
58
59
Головка нижней челюсти. У новорожденного эта головка округлой формы и имеет почти одинаковые поперечный (медиолатеральный) и переднезадний размеры. С возрастом она постепенно удлиняется в попереч-ном направлении. С момента прорезывания молочных зубов и до двух лет происходит увеличение головки. После этого наступает стабилизация размеров головки, которая длится до шести лет, когда появляется первый постоянный зуб, после чего вновь размеры головки увеличиваются. У новорожденного еще не выражен наклон головки кпереди. С возрастом головка наклоняется кпереди по отношению к шейке суставного отростка. В грудном возрасте нижняя челюсть занимает дистальное положение. С прорезыванием молочных моляров и увеличением высоты прикуса происходит дальнейшее перемещение суставной головки кпереди. В передне-верхнем отделе суставной головки находится суставная поверхность, покрытая хрящом. У новорожденного головка покрыта толстым слоем волокнистой соединительной ткани, а у взрослых — волокнистым хрящом, который с возрастом истончается.
Головка взрослого имеет эллипсоидную форму, она вытянута в поперечном направлении и сдавлена в переднезаднем направлении, ее длинная (медиолатеральная) ось примерно в 3 раза больше, чем переднезадняя. Обе головки челюсти не стоят строго во фронтальной плоскости, а их горизонтальные длинные оси сводятся под углом, открытом кпереди, и совпадают с поперечным диаметром нижнечелюстных ямок. Головка состоит из тонкого слоя компактной кости, под которым находится губчатое вещество.
Шейка нижней челюсти сужена, на ее передней поверхности находится крыловидная ямка, где прикрепляется большая часть верхней головки латеральной крыловидной мышцы. Формирование крыловидной ямки наблюдается в возрасте 5 лет и имеет вид узкой неглубокой поперечной бороздки. В норме суставная головка передает давление через бессосудистую центральную часть внутрисуставного диска на задний скат суставного бугорка.
Нижнечелюстная ямка. Служит вместилищем для головки нижней челюсти. У новорожденного она почти плоская, округлой формы. Спереди она не ограничена суставным бугорком, а сзади имеется хорошо выраженный суставной конус. Последний предохраняет барабанную часть среднего уха от давления суставной головки. По мере развития суставного оу-горка позадисуставной конус атрофируется. У новорожденного нижнечелюстная ямка функционирует полностью, так как нижняя челюсть смешена дистальне и суставная головка располагается в заднем ее отделе. Толщина кости свода ямки у новорожденного несколько превышает 2 мм В дальнейшем глубина нижнечелюстной ямки увеличивается. Это связано с
ростом скулового отростка височной кости, который формирует суставной бугорок и обеспечивает углубление суставной ямки и отделение суставной поверхности от височной поверхности чешуи. С возрастом суставная ямка увеличивается преимущественно в поперечном направлении и углубляется, что соответствует изменениям головки нижней челюсти и имеет эллипсоидную форму. Суставная поверхность покрыта волокнистым хрящом.
Поперек нижнечелюстной ямки, примерно в дистальной трети, ее пересекает каменисто-барабанноя (глазерова) щель и делит ямку на переднюю — интракапсупярную часть (лежащую в полости сустава) и заднюю — экстракапсулярную часть (лежащую вне полости сустава). Поэтому интракапсулярная часть называется суставной ямкой.
Размеры нижнечелюстной ямки в 2—3 раза больше головки нижней челюсти, поэтому имеет место инконтруэнтность (несоответствие размеров головки и ямки). Неконгруэнтность сочленяющихся поверхностей сустава выравнивается благодаря сужению размеров ямки за счет прикрепления суставной капсулы внутри нее у переднего края каменисто-барабанной щели височной кости, а также компенсируется суставным диском, делящим полость сустава на две камеры, обеспечивая высокую конгруэнтность суставных поверхностей. Суставной диск прилегает к суставным поверхностям и повторяет форму головки нижней челюсти и заднего ската суставного бугорка, увеличивая площадь соприкосновения суставных поверхностей.
Суставной бугорок. У новорожденного суставной бугорок отсутствует, он только намечается впереди нижнечелюстной ямки. С ростом основания скулового отростка височной кости и прорезыванием молочных зубов размеры суставного бугорка постепенно увеличиваются. В возрасте 6 7 лет он уже хорошо заметен. Суставной бугорок у взрослого представляет собой эллипсоидное костное возвышение в форме цилиндра височной кости, лежащее поперечно в заднем отделе скулового отростка височной кости, длинная ось которого направлена так же, как и у нижнечелюстной ямки. Он имеет передний скат, гребень (вершину) и задний скат. Суставными поверхностями являются гребень и задний скат, которые покрыты волокнистым хрящом.
Внутрисуставной диск. Повторяет формы сочленяющихся поверхностей и располагается между ними. У новорожденного суставной диск представляет собой мягкую прослойку округлой формы, вогнутую снизу и выпуклую сверху с едва заметными утолщениями спереди и сзади. Состоит из коллагеновых волокон. По мере того, как формируются костные образования сустава, параллельно формируется и диск. Такие изменения с диском направлены на обеспечение конгруэнтности суставных поверхно-
60
61
стей. Внутрисуставной диск постепенно приобретает переднее и заднее утолщение и тонкую центральную часть. Верхняя височная поверхность диска выпуклая сзади и седлообразная спереди, а нижняя вогнута — повторяет форму головки нижней челюсти и создает как бы дополнительную подвижную ямку.
Выделяют четыре зоны диска (рис. 26):
передний полюс диска;
промежуточная зона — средняя часть, самая тонкая часть с хорошей эластичностью и гибкостью;
задний полюс диска — толще и шире переднего;
биламинарная зона («задисковая подушка») — располагается между задним полюсом диска и капсулой сустава, представлена двумя связками, между которыми расположена нервно-сосудистая зона.
сустава, позволяя диску и головке совершать небольшие переднезадние движения вокруг вертикальной оси.
Диск занимает такое положение в полости сустава, что при движении головки нижней челюсти наибольшее давление приходится на задний скат и вершину суставного бугорка, а не на тонкую костную пластинку верхней и задней части нижнечелюстной ямки. Тем самым диск является мягкой и упругой прокладкой, амортизирующей силу жевательного давления. Внутрисуставные связки. Прикрепление диска показано на рис. 27.
Центральная часть диска является областью вращения, в ней нет сосудов и нервов. Диск по краям сращен с капсулой сустава на всем протяжении и делит полость сустава на два несообщающихся между собой отдела. Верхний отдел расположен между верхней поверхностью диска и суставными ямкой и бугорком. Нижний отдел сустава образован головкой нижней челюсти и нижней поверхностью диска.
Верхний отдел сустава с медиальной и латеральной стороны образует карманы у полюсов головки нижней челюсти между диском и капсулой сустава. На дне этих карманов находятся медиальная и латеральная диско-челюстные связки, идущие от суживающихся боковых краев диска к медиальному и латеральному полюсам суставной головки и прикрепляющиеся позади и ниже последних подобно чепчику, сидящему на голове Это сращение образует своеобразную ось вращения для нижнего отдела
Спереди передний полюс диска соединяется следующим образом. Верхняя часть диска соединяется с височной костью передней дисковисочной связкой. Нижняя часть диска соединяется с головкой нижней челюсти передней дискочелюстной связкой. Они имеют прямоугольную форму. Соединение переднего полюса диска с капсулой сустава имеет очень важное значение в понимании внутрисуставных изменений. С внешней стороны капсулы в ее переднемедиальную поверхность вплетаются волокна верхней головки латеральной крыловидной мышцы. Некоторая часть этих волокон непосредственно прикрепляется к переднемеди-альной поверхности внутрисуставного диска.
Задняя зона прикрепления диска — биламинарная зона — представлена двумя связками. Верхняя связка состоит из эластина и прикрепляется сзади к барабанной части височной кости, это — задняя дисковисочная связка. При смещении суставной головки и диска вперед она натягивается
62
63
и действует как сила, противоположная силе сокращения латеральной крыловидной мышцы, а при закрывании рта возвращает мениск в исход, ное положение. Нижняя связка состоит из коллагена и прикрепляется сзади и снизу суставной головки — задняя дискочелюстняя связка. При смещении суставной головки и диска вперед она смещается вперед вместе с ними до определенного состояния, после чего препятствует этому смещению.
Между верхним и нижним слоями биламинарной зоны находится богатая сосудами и нервами зона. На сагиттальном разрезе биламинарная зона имеет форму трапеции, большее основание которой находится у капсулы сустава, а меньшее — у суставного диска. При смещении головки вместе с диском вперед биламинарная зона наполняется кровью, тем самым заполняя освобожденное головкой пространство. По мере того, как происходит возвращение головки с диском в исходное состояние, биламинарная зона сжимается и освобождается от крови. Эту периодичность называют физиологическим процессом гемодинамики.
Суставная капсула. Она определяет анатомические и физиологические пределы ВНЧС. Суставная капсула представляет собой эластичный соединительнотканный «мешок», в который заключены суставные поверхности сочленяющихся костей, и соединяется с диском по его периметру. Имеет вид «воронки», суживающейся книзу. Прикрепление капсулы к височной кости как бы сдвинуто кпереди по отношению к нижнечелюстной ямке. Сзади она прикрепляется вдоль переднего края каменисто-барабанной (глазеровой) щели и делит нижнечелюстную ямку на переднюю внутрикапсулярную и заднюю внекапсулярную части. Капсула также окружает суставную поверхность головки нижней челюсти. Характеризуется высокой прочностью и эластичностью и не рвется при полных вывихах сустава.
Состоит из двух слоев: наружного, представленного фиброзной соединительной тканью, и внутреннего — эндотелиалъного (синовиальный слой). Клетки синовиальной оболочки вырабатывают синовиальную жидкость, являющуюся основным субстратом, обеспечивающим трофику суставного хряща.
Синовиальная жидкость. Функции синовиальной жидкости:
локомоторная — обеспечивает свободное скольжение суставных поверхностей;
метаболическая — принимает участие в процессе обмена между полостями сустава и сосудами, а также в перемещении и ферментативном распаде клеток с последующим удалением их из полости сустава по лимфатическому руслу;
трофическая — осуществляет питание бессосудистых слоев суставного диска, суставных поверхностей и других элементов сустава;
— защитная — принимает участие в ликвидации чужеродных клеток и веществ, проникающих из крови, при повреждении суставной капсулы и др.
Синовиальная оболочка образует складки в передней и задней поверхности сустава. В зависимости от движения вперед или назад складки расправляются. Так, при движении головки и диска вперед складки образуются спереди, а сзади расправляются. При движении головки и диска назад —наоборот.
В области биламинарной зоны клетки синовиальной оболочки образуют выросты, так называемые ворсины, которые являются участками интерорецепции. В зависимости от возраста количество и расположение их различно. У новорожденного ворсины отсутствуют. Небольшое их количество появляется в возрасте 1—2 лет и увеличивается к 3—6 годам жизни ребенка. В 16—18 лет их уже большое количество. По мере старения организма идет инволюция ворсин.
Капсулу сустава со всех сторон усиливают связки. Связки делятся на внутри- и внекапсулярные.
Внутрикапсулярные связки находятся внутри сустава. Их шесть: передняя, задняя, латеральная и медиальная дискочелюстные; передняя и задняя дисковисочные. Они описаны выше.
Внекапсулярные связки. Наиболее прочной из внекапсулярных связок является латеральная связка. Она прилежит к капсуле сустава и сплетается с ней на ее латеральной поверхности (рис. 28, а). Связка берет начало от задней части скулового отростка височной кости латеральнее суставного отростка и косо веерообразно идет назад и книзу (сужаясь), прикрепляясь ниже и позади латерального полюса суставной головки. На своем пути она отдает горизонтальные глубокие волокна к капсуле. Главная биомеханическая функция этой связки — приостанавливать или ограничивать движения комплекса головка—диск и ограничивать смещение нижней челюсти назад к позадимыщелковым структурам биламинарной зоны. Она также регулирует боковые и сагиттальные движения нижней челюсти. Это наиболее важная связка.
Клиновидно-нижнечелюстная связка (рис. 28, б) несколько отстоит от медиальной поверхности капсулы, начинаясь от угловой ости клиновидной кости и прикрепляясь к язычку нижней челюсти. Ограничивает боковые и задние смещения нижней челюсти.
Шилонижнечелюстная связка далеко отстоит от сустава, начинается от шиловидного отростка и прикрепляется к углу нижней челюсти. Ограничивает смещение нижней челюсти вперед.
64
65
Ниже представлен механизм суставных изменений, который позволяет нижней челюсти совершать полный комплекс свойственных ей движений.
При вертикальных движениях (открывание рта) (рис. 29) в начальной фазе головка вращается вокруг горизонтальной оси в нижнем отделе сустава (при открывании рта до 2 см). Затем эти движения сочетаются с поступательными в верхнем отделе, где суставные головки вместе с дисками начинают выдвигаться вперед и вниз, скользя по заднему скату суставного бугорка (открывание рта до 5 см). В конце пути, когда головки достигают крайнего положения, вновь происходят только вращательные движения вокруг горизонтальной оси в нижнем отделе.
Связки состоят из фиброзной неэластичной соединительной ткани, что препятствует растяжению капсулы сустава при нормальном объеме движений нижней челюсти. В случае перерастяжения связок первоначальная длина их не восстанавливается.
ВНЧС имеет очень сложную систему иннервации и кровоснабжения.
Иннервация ВНЧС. Иннервация сустава осуществляется различными нервами. Передняя часть сустава иннервируется жевательным, задним глубоким височным и латеральным крыловидным нервами. Наружную часть иннервируют жевательный и ушно-височный нервы. Внутренняя и задняя поверхности иннервируются ушно-височным нервом. От перива-скулярных сплетений отходят веточки, участвующие в иннервации сустава.
Кровоснабжение ВНЧС. Основными источниками кровоснабжения сустава являются две магистральные артерии (верхнечелюстная и поверхностная височная) и их многочисленные ветви.
Биомеханика височно-нижнечелюстного сустава
Движения в ВНЧС у новорожденного и взрослого человека различны С момента рождения и до 7—8 мес. жизни ребенка доминируют сагиттальные движения нижней челюсти, связанные с актом сосания. Такой характер движений в ВНЧС обусловлен строением его у новорожденного и обеспечивается скольжением округлой суставной головки вместе с диском по достаточно плоской ямке. По мере прорезывания молочных зубов и развития суставных бугорков появляются откусывающие, разжевывающие, боковые движения нижней челюсти.
Выдвижение нижней челюсти вперед (сагиттальные движения) при сомкнутых зубах из положения центральной окклюзии в переднюю в большинстве случаев направляется поверхностями смыкания передних зубов. Во время сагиттальных движений головки перемещаются вниз и вперед вдоль скатов суставных бугорков. При движении вниз головки также совершают вращательные движения в нижнем отделе сустава, заставляя нижнюю челюсть совершать открывающие движения, диктуемые направляющими скатами передних зубов (рис.30).
Способность головок перемещаться вперед вместе с диском по суставным скатам и одновременно вращаться в нижнем отделе позволяет нижней челюсти следовать сагиттальному резцовому пути (это путь, который проходят нижние резцы по небным поверхностям верхних резцов при движении нижней челюсти из центральной окклюзии в переднюю), в то время как задние зубы разомкнуты (дезокклюзия). В конце сагиттального суставного пути (это путь, который проходят головки вниз и вперед по заднему скату суставного бугорка), при движении из передней окклюзии в крайнее переднее положение к поступательным движениям в верхнем отделе присоединяются вращательные движения вокруг горизонтальной
66
67
70. Понятие иммунитета. Неспецифические факторы защиты
Иммунитет – способность поддерживать генеостаз (постоянство внутренней среды). Все средства защиты разделяются на специфические и неспецифические. Специфические - появляется иммунитет через 48 часов после контакта с патогенном (латентный период) и действует против строго определённого патогенна – адентивный иммунный ответ. Неспецифические факторы защиты препятствуют размножению патогена в латентный период; действуют против любого патогенна с разной эффективностью – воспалительная реакция. К ним относят кожа и слизистая, клеточная защита, гуморальная защита. Кожа и слизистая: явл-ся преградой на пути микробов; на поверхности кожи высокое осмотическое давление, молочная кислота, ненасыщенные жирные кислоты; слизистая выделяет секреты с бактерицидными свойствами, в том числе желудочный и кишечный сок. Всё это неблагоприятно для развития микробов. Если микробы внедрились в подкожную клетчатку или в подслизистую ткань, место внедрения – ворота инфекции. Сюда устремляется группа фагоцитов. Клеточная защита – фагоцитоз. Гл роль в нем играют лейкоциты – при остром воспалении; фагоциты – при хроническом воспалении. Микроб, захваченный фагоцитом, может подвергнуться полному перевариванию – завершённый фагоцитоз. Микроб внутри лейкоцита размножается – незавершённый фагоцитоз. В таком виде патоген не доступен действию антител. Многие микробы имеют капсулы, выделяют токсины => полиинфекции покрываются погибшими эритроцитами микробов и продуктами распада. Развивается воспаление. В этот очаг поступает жидкая часть крови и лимфы, кот содерж гуморальные факторы защиты – стволовые лимфоидные клетки превращаются в В – лимфоциты, кот ответственны за реализацию гуморального иммунного ответа. В – система ответственна за иммунитет при многих бактериальных инфекциях, антитоксический иммунитет, аллергию немедленного типа. В – лимфоциты имеют рецепторы – макромолекулярные структуры клеточной поверхности, с помощью кот клетки узнают антигены.
72. Лимфоциты: т и в – типа. Их функции
Лимфоциты содержат крупное ядро, окружённым узким ободком слабо базафильной цитоплазмы, органоиды слабо развиты. По функционному признаку различают: Т – лимфоциты проходят развитие в вилочковой железе и в спец зонах переферич лимфоидных органоидов. Долгоживущие. Обеспечивают реакции клеточного иммунитета, участвуют в гуморальном иммунитете. Среди них различают: Т – клетки памяти – долго живут, сохраняя информацию об антигене, кот вызвал их появление. Т –киллеры – обладают цитотоксичным эффектом по отношению к чужеродным клеткам. Т- хелперы – помощники – в гуморальном иммунитете – помогают В- лимфоцитам вырабатывать иммуноглобулин. Т – супрессоры – подавляют способность В - лимфоцитов вырабатывать иммуноглобулин. В-лимфоциты развиваются в красном костном мозге и переферич лимфоидных органах. Короткоживущие. В – клетки памяти – сохраняют информацию об антигене.
73. Иммунный ответ. Локус иммунного ответа
Иммунный ответ, или иммунологическая реактивность, - высокоспецифическая форма реакции организма на чужеродные вещества (антигены). При иммунном ответе происходят распознавание чужеродного агента. При введении антигена возникает первичный иммунный ответ - через 2 дня в крови образуются антитела, титр которых возрастает, достигает максимума, а затем падает. Вторичный иммунный ответ возникает на повторное введение того же антигена и характеризуется более высоким и быстрым нарастанием титра антител. Подобная реакция более усиленного образования антител на повторное введение антигена – иммунологическая память При вирусной инфекции ДНК или РНК вируса попадает в клетку, а вирусные белки остаются на клеточной мембране. Цитотоксические Т-киллеры своими рецепторами узнают вирусные антигены только в комбинации с белком главного комплекса гистосовместимости МНС класса 1.В отличие от антител Т-рецепторы не узнают и не связывают антиген, если тот не находится вместе с белком МНС. После узнавания антигенов цитотоксические Т-клетки убивают зараженные вирусом клетки. Мутации любых локусов, обусловливающие разные звенья иммунной системы организма, влияют на иммунный ответ. Гены иммунного ответа. Гены, кодирующие иммунный ответ, наз-ся генами иммунного ответа Высота иммунного ответа детерминирована многими генами иммунного ответа, обозначаемыми Iг-1, Iг-2 и т. д. Контроль иммунного ответа осуществляется Iг-генами путем контроля синтеза Iа-белков. Во многих случаях иммунный ответ против антигенов наследуется полигенно. Гены иммунного ответа: 1) Ir-гены определяют количество синтезируемых антител против определенных антигенов; 2) Ir-гены не сцеплены с локусами, кодирующими иммуноглобулины; 3) Ir-гены высокоспецифичны. 4) между генами, контролирующими высокий или низкий иммунный ответ против различных антигенов, в основном не существует никакой связи. Теории иммунитета: 1) клонально-селекционная теория Ф. Бернета (1959). Она основана на четырех основных принципах: а) в организме имеется большое число лимфоидных клеток; б) популяция лимфоидных клеток гетерогенна, и в результате интенсивного деления клеток образуется большое число клонов; в) небольшое количество антигена стимулирует клон клеток к размножению; г) большое количество антигена элиминирует соответствующий клон. 2) Сетевая теория. Согласно неё антитела не только узнают антиген, но и сами являются антигенами.