Сорусы папоротника это
Сорус — Википедия
Буро-красные точки — сорусы на нижней стороне вайи папоротникаСо́рус (от др.-греч. σορός — погребальная урна) — группа расположенных скученно спор или органов бесполого размножения — спорангиев или гаметангиев на поверхности таллома у красных и бурых водорослей, на листьях у папоротниковидных, а также группа плодовых тел у низших грибов.
- Медиафайлы по теме сорус в Викискладе
- Сорусы папоротников на сайте Биология и медицина (Проверено 1 ноября 2009)
Для улучшения этой статьи желательно:
|
Папоротники, подготовка к ЕГЭ по биологии
Папоротники относятся к наиболее древним споровым растениям. Обитают в самых разнообразных экологических условиях: в заболоченных местах и водоемах, лесах тропического и умеренного климата. Наиболее известные представители - щитовник мужской, орляк, страусник. В жизненном цикле папоротников, хвощей, плаунов доминирует спорофит.
Расцвет папоротников произошел около 358 млн лет назад и длился примерно 65 млн лет, в честь папоротников назван период палеозойской эры - карбон, или каменноугольный период, который длился указанный промежуток времени. Именно папоротникам принадлежит активная роль в углеобразовании: в карбоне леса заселяли древовидные папоротники, достигавшие 40 и более метров в высоту.

Древесные формы папоротников сохранились до наших дней, однако большинство представителей - травянистые растения, у которых отсутствует камбий, а значит, нет и вторичной древесины.
Папоротники относятся к группе сосудистых растений, так как имеют жилки - сосудисто-волокнистые пучки, в отличие от мхов, которые не имеют жилок и не являются сосудистыми растениями. Механическая прочность обеспечивается отложением склеренхимы вокруг проводящих пучков (жилок).

В отличие от мхов, папоротники имеют проводящую ткань в стеблях и корнях, состоящую из ксилемы и флоэмы. Заметьте, я написал "корень" - у мхов не было и корней, вместо них были ризоиды, выполняющие аналогичную корням функцию. Корни папоротников, хвощей и плаунов всегда придаточные и растут от видоизмененного побега - корневища.
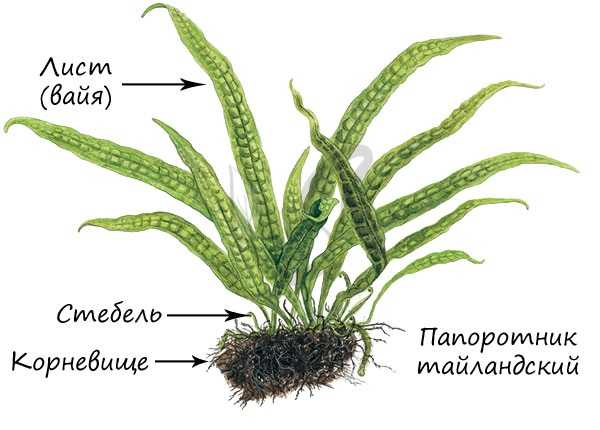
Строение
Рассмотрим строение папоротников на примере типичного представителя - щитовника мужского. Это широко распространенный папоротник, типичный для климата умеренных широт. Представляет собой многолетнее травянистое корневищное растение.
Образован пучком сильно расчлененных листьев, отходящих от корневища. Нарастают листья верхушкой, образуя завитки - "улитки".

Отметьте, что лист папоротника называется вайя (от греч. baion - пальмовая ветвь). В отличие от настоящих листьев, вайя имеет недетерминированный верхушечный рост. Листья имеют черешок, прикрепляющийся к стеблю, который может продолжаться в рахис - главную ось сложного листа, соответствующую центральной жилке.

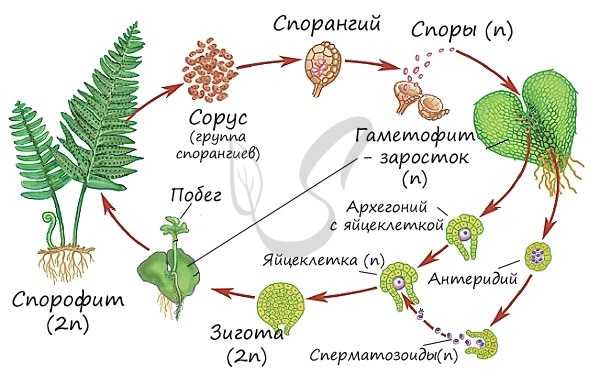
Жизненный цикл папоротниковидных
Листостебельное растение папоротника, изображенное выше - спорофит (2n). Спорофит доминирует в жизненном цикле папоротниковидных, в отличие от цикла мха, где спорофит, по сути, является придатком гаметофита (редуцирован). На нижней стороне вайи располагаются спорангии, собирающиеся в сорусы - группы близкорасположенных спорангиев. На спорофите (2n) в спорангии после мейоза образуются споры (n).

В 2012 году группа ученых под руководством Ксавьера Нобли из Университета Ниццы выяснили, что спорангий имеет специальный механизм "катапульты", споры из него вылетают со скоростью около 10 м/с.
Гаплоидные споры (n) прорастают в заросток (n), небольшую пластинку (несколько мм) сердцевидной формы. Заросток зеленого цвета, способен фотосинтезировать и прикрепляется к почве ризоидами. На нем образуются мужские и женские половые органы - соответственно антеридии и архегонии. Сперматозоид (n), образовавшийся в антеридии, благодаря воде (во время дождя) попадает в архегоний, где сливается с яйцеклеткой (n) и образуется зигота (2n).
Из зиготы развивается зародыш, который проникает в ткани архегонии с помощью особого приспособления - гаустории (от лат. haustor - черпающий, пьющий). Гаустория представляет собой ножку, внедряющуюся в ткани заростка и поглощающую из него питательные вещества. Начинается бурный рост зародыша, образуется побег, а затем взрослое растение - спорофит (2n). Цикл замыкается.
Значение папоротников
Папоротники являются главным компонентом многих лесных сообществ, звеном в цепи питания - продуцентами (производителями органических веществ). Человек использует папоротник в декоративных целях. Молодые побеги некоторых папоротников съедобны и употребляются в пищу: побеги орляка, листья страусника обыкновенного.
Щитовник мужской имеет медицинское значение: из его корневищ изготавливают противогельминтный препарат.
© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Спорангии и споры - это... Что такое Спорангии и споры?
На определенной стадии развития спорофита начинается его спороношение — явление, в биологическом отношении аналогичное цветению. Внутренние физиологические процессы, вызывающие переход спорофита из вегетативной фазы развития в фазу спороношения, еще далеко не разгаданы, но, вероятно, они имеют мпого общего с теми процессами, которые приводят к цветению. Спороношение начинается с закладки тех начальных (ипициальпых) клеток или групп клеток, из которых развивается спорангий. Как мы уже знаем, в одних случаях спорангии развиваются на обыкновенных зеленых листьях, в других случаях — на специальных спороносящих частях листа или на особых листьях, резко отличающихся от вегетативных и специализированных для функции спороношения. Обособление фертильных участков и специализированных фертильных листьев (спорофиллов) — явление в эволюционном смысле вторичное.
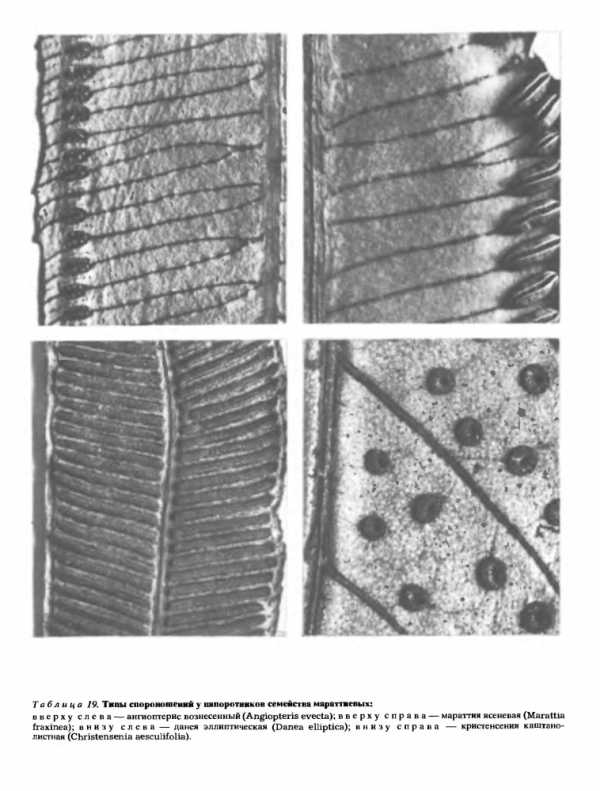
,

Интересно, что, в то время как у ужовниковых и мараттиевых (Marattiaceae) каждый спорангий развивается из группы начальных клеток и имеет многослойную стенку, почти у всех остальных современных папоротников он развивается из одной-единственной начальной клетки и имеет однослойную стенку (табл. 19, 20). Очевидно, при втором способе развития спорангия требуется меньше строительного материала (меньше клеток), а само развитие происходит быстрее. Поэтому все согласны с тем, что возникновение спорангиев, берущих начало от одной клетки и имеющих однослойную стенку, представляет в эволюционном отношении шаг вперед.
Вспомним еще раз, что листья папоротников возникли в процессе эволюции из ветвей риниофитовых предков. Отсюда вполне понятно, что у древнейших папоротников спорангии имели верхушечное (терминальное) расположение, т. е. сидели на верхушках конечных веточек дихотомически разветвленного листа. Среди ныне живущих папоротников расположение спорангиев, наиболее близкое к верхушечному, наблюдается у гроздовника (Botrychium), относящегося к примитивному семейству ужовниковых.
По мере того как примитивный, похожий еще на ветку лист уплощается, между отдельными веточками образуется как бы перепонка (подобно плавательной перепонке водоплавающих птиц), представляющая собой результат бокового разрастания ткани веточек. Образование перепонки увеличило фотосинтезирующую поверхность листа, что дало огромное преимущество таким растениям. Спорангии оста лись, в сущности, на своих прежних местах. Но так как верхушки веточек оказались у нового органа растения — листа — концами жилок (ведь каждая веточка жилкования соответствует первоначальному проводящему пучку веточки), спорангии автоматически оказываются на краю пластинки листа. Таким образом верхушечное расположение спорангиев переходит в краевое (маргинальное). Трудно сказать, было ли это на самом деле шагом вперед или только естественным результатом возникновения листовой пластинки. Во всяком случае, краевое расположение спорангиев наблюдается у многих современных папоротников.
Значительным шагом вперед в приспособительной эволюции папоротников был переход спорангиев из краевого расположения в поверхностное (ламинальное). Этот процесс происходил независимо в рззных линиях эволюции, причем во всех случаях спорангии перемещались на нижнюю, или «брюшную» (абаксиальную), сторону листовой пластинки. Это вполне понятно, так как на брюшной стороне они больше защищены от всяких неблагоприятных воздействий внешней среды и находятся в лучших условиях питания и рассеивания зрелых спор. Кроме того, если бы спорангии перешли на верхнюю, или «спинную» (адаксиальную), сторону пластинки, они бы заняли некоторую часть наиболее активной фотосинтезирующей поверхности, что неизбежно отразилось бы на эффективности фотосинтеза. Поэтому не удивительно, что во всех без исключения случаях поверхностного расположения спорангиев они занимают нижнюю сторону листовой пластинки. У ныне живущих папоротников наблюдаются все переходные стадии от краевого расположения к поверхностному (а последнее встречается чаще других). Более того, у некоторых из тех папоротников, спорангии которых сидят вдоль края пластинки, зто перемещение из краевого положения в поверхностное осуществляется в процессе онтогенеза.
Наряду с перемещением спорангиев на нижнюю сторону пластинки происходит еще и другое, не менее важное изменение в их расположении. Сначала спорангии бывают расположены одиночно, т. е. не образуют групп, как, например, у схизейных. Но уже у некоторых папоротников с краевым расположением, как, например, у диксонии (Dicksonia) и локсомы (Loxsoma) или у всех гименофилловых, спорангии сгруппированы в отдельные спорангиальные кучки, или сорусы (от греч. soros — куча). У папоротников с поверхностным расположением спорангиев это уже становится правилом. Возникновение сорусов связано прежде всего с увеличением числа спорангиев. Так, если у предков диксонии на конце жилки сидело лишь по одному спорангию, то у самой диксонии — целая спорангиальная кучка. Но не менее важно и то, что с образованием сорусов возникает возможность более концентрированного и, следовательно, более эффективного снабжения спорангиев питательными веществами.
В биологическом отношении чрезвычайно важен порядок развития и созревания спорангиев на листе или в каждом отдельном сорусе. У вымерших папоротниковидных и у ныне живущих ужовниковых, мараттиевых, осмундовых, плагиогириевых, схизейных, глейхениевых и матонневых все более или менее близко расположенные на растении спорангии или все спорангии внутри соруса (плагиогириевые, глейхениевые и матониевые) развиваются одновременно (симультанно). Сорусы, в которых развитие и созревание спорангиев происходит одновременно, называют простыми.
Одновременное развитие спорангиев имеет один существенный недостаток. Дело в том, что при ухудшении условий произрастания и физиологическом угнетении растения подвергается риску развитие одновременно формирующихся спорангиев на части растения или даже на всем растении. Выходом из этого положения является последовательное развитие спорангиев, когда их созревание в сорусе наступает в разное время. Это достигается посредством удлинения ложа (рецептакула) соруса и увеличения количества спорангиев при одновременном уменьшении их размеров. Устанавливается так называемая базипетальная (от греч. basis — основание и лат. реtere — устремляться) последовательность, когда развитие начинается с верхней части ложа (в центре соруса) и постепенно переходит к основанию ложа (к периферии соруса). В результате самые молодые спорангии оказываются наиболее близко расположенными к источнику снабжения питательными веществами и наиболее защищенными. Все развитие соруса растягивается во времени, и поэтому процесс спороношения оказывается сравнительно менее уязвимым (если пострадают более ранние спорангии, то будет еще шанс для развития более поздних). Особенно хорошо выражено базипетальное развитие спорангиев у представителей семейства гименофилловых, имеющих длинное цилиндрическое или линейное ложе. Сорусы с базипетальным развитием спорангиев называют градатными (от лат. gradatio — постепенное возвышение, усиление). Такие сорусы характерны для циатейных, гименофилловых, аснлениевых, а также для всех разноспоровых папоротников.
В некоторых линиях эволюции папоротников из градатных или прямо из простых сорусов возникают так называемые смешанные сорусы, характеризующиеся неопределенной последовательностью в развитии спорангиев. В таких сорусах молодые и более зрелые спорангии расположены вперемешку, без определенного порядка. Несмотря на то что ложе таких сорусов обычно плоское, спорангии имеют разную высоту, так как у более зрелых спорангиев ножка удлиняется, и поэтому они заметно возвышаются над более молодыми. Преимущество смешанных сорусов заключается прежде всего в том, что развитие спорангиев в сорусе растягивается во времени. Но не менее важно то обстоятельство, что молодые спорангии здесь надежно защищены возвышающимися над ними зрелыми спорапгиями, оставшимися ножками уже раскрывшихся спорангиев, а также волосками, которые часто сидят на ложе между спорангиями. Исключительно важно также, что в смешанных сорусах может эффективно и без всяких физических помех действовать наиболее совершенный способ раскрывания спорангиев.
Смешанные сорусы возникают независимо в разных линиях эволюции папоротников, притом они происходят как от простых, так и от градатных сорусов. Происхождение от простых сорусов можно наблюдать в пределах рода диптерис, где у одного из видов (диптерис Лобби — Dipteris lobbiana) развитие спорангиев симультанное, в то время как у более подвинутого вида — диптериса двойчатосложного (D. conjugate), развитие спорангиев происходит в неопределенной последовательности (сорусы смешанные). У близкого к диптерисовым и, вероятно, производного от него семейства хейроплевриевых (Cheiropleuriaceae) сорусы также смешанные. У большого и широко распространенного семейства полиподиевых, имеющего общее происхождение с двумя выше упомянутыми маленькими семействами, также установилась неопределенная последовательность в развитии спорангиев. Таков же порядок развития у другого большого семейства, адиаптовых.
В эволюции сорусов произошло еще одно важное событие. Речь идет о защитных приспособлениях спорангиев. Вообще защита молодых спорангиев у папоротников обеспечивается хотя бы тем, что их молодые листья скручены спирально как раковина улитки, причем не только по главной жилке, но и по всем сегментам. Это проявилось уже у некоторых вымерших примитивных родов. Нередко перышки остаются еще закрученными, когда большая часть листа уже расправилась. Кроме этой общей защиты, почти универсальной для папоротников (за исключением ужовниковых), имеются также более специальные формы защиты, обеспечиваемые различными типами п окрывальца, или индузия (от лат. indusium — верхняя туника). Термин этот служит для обозначения «покрывалец» самого различного происхождения. Покрывальца имеются далеко не всегда. Простые сорусы с характерным для них одновременным развитием спорангиев нуждаются в покрывальце в меньшей степени, чем два других типа сорусов, у которых развитие спорангиев более растянуто. Поэтому у большинства семейств с простыми сорусами покрывальце совершенно отсутствует. Однако у схизейных отдельные спорангии (сорусов у них нет) защищены загнутыми на нижнюю сторону краями пластинки листа, или же у видов лигодиума (Lygodium) каждый спорангий полностью прикрыт особым кармашковидным выростом поверхностных тканей листа. Имеются и другие формы защиты, в том числе взаимная защита спорангиев в результате их срастания в так называемые синангии (например, у некоторых мараттиевых). Настоящее же покрывальце среди папоротников с простыми сорусами имеется только у матонии (Matonia). Оно очень своеобразное, зонтиковидной формы и возникает из центральной части ложа соруса. По всем данным, оно произошло совершенно независимо от покрывалец градатных и смешанных сорусов, к которым мы теперь перейдем.
Градатные сорусы, характеризующиеся более длительным развитием и более мелкими спорангиями, нуждаются в защите гораздо больше, чем простые сорусы с более крупными и одновременно развивающимися спорангиями. Поэтому в разных линиях эволюции папоротников с градатными сорусами независимо и разными путями возникли покрывальца, эффективно защищающие развивающиеся спорангии. Покрывальца бывают различной формы, чаще всего более или менее чашевидные или двугубые. Приспособительная эволюция покрывалец представляет собой одну из наиболее увлекательных глав сравнительной морфологии папоротников, но в ней еще много неясного. В частности, не всегда понятно исчезновение в процессе эволюции уже имевшегося у предков покрывальца. Что же касается папоротников со смешанными сорусами, то за исключением большинства даваллиевых (Davalliaceae) они лишены покрывальца. Функцию покрывальца часто исполняют у них перемешанные со спорангиями волоски (парафизы).
Вернемся снова к спорангию. При созревании он раскрывается, а споры освобождаются и рассеиваются. Нас интересуют, однако, те приспособления, которые способствуют раскрыванию спорангия. Начпем с самых древних, вымерших форм. У некоторых из них спорангий при созревании открывался наверху небольшим отверстием. У пих, так же как у современных ужовниковых (обладающих верхушечным щелевидным отверстием), нет еще никаких дополнительных приспособлений, способствующих раскрыванию зрелого спорангия. Но уже у целого ряда древнейших папоротниковидных, например у представителей палеозойского семейства зигоптерисовых (Zygopteridaceae), спорангии снабжены с обеих сторон широкой продольной (вертикальной) полоской из нескольких рядов клеток с утолщенными стенками, служившей для раскрывания спорангиев. Это чисто механическое приспособление называют кольцом (рис. 91). Мы не знаем, как действовало у вымерших форм кольцо, по его механизм у современных папоротпиков хорошо изучен.
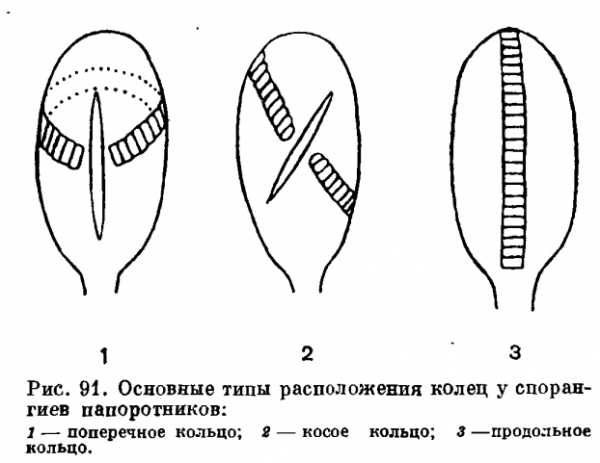
Познакомимся поэтому со строением и действием наиболее распространенного типа кольца ныне живущих папоротников. Это продольное (вертикальное) образование, состоящее из одного ряда клеток. У большей части клеток кольца три внутренние стенки (две радиальные и внутренняя тапгентальная стенка) утолщены, в то время как остальные три (внешние тангептальные стенки) тонкие. Одним своим концом эта часть кольца с утолщенными стенками клеток прочно прикреплепа к пожке спорангия. Часть клеток кольца (четверть или треть от общего их количества) имеет относительно тонкие стенки, включая группу из четырех поперечно удлиненных сестринских клеток, образующих так называемый стомий (от греч. stoma — рот). По мере созревания спорангия клетки кольца в результате испарепия постепеппо теряют воду. Но дело в том, что имеется очень сильное «сцепление» между стенками клеток кольца и водой. Поэтому, когда через тонкие внешние тангепталыше стенки вода испаряется, эти стенки благодаря силам сцепления постепенно втягиваются внутрь и становятся все более вогнутыми, в то время как концы радиальных клеток стягиваются друг к другу. По мере того как стенки клеток кольца деформируются, возникает очень сильное механическое напряжение, равное давлению около 300 или более атмосфер. В результате возрастающего напряжения в конце концов наступает момент, когда стенка спорангия разрывается в районе стомия, а само кольцо медленно загибается в обратную сторону, как бы выворачиваясь наизнанку. По мере того как вода продолжает испаряться, возрастающее напряжение деформированных стенок преодолевает силы сцепления и внезапно и одновременно во многих клетках кольца образуется вакуум. Силы сцепления таким образом уже перестают действовать, и кольцо быстро и с силой возвращается к своему исходному положению. В этот момент оно действует подобно катапульте и с силой выбрасывает освободившиеся уже споры. Таким образом, перед нами удивительно остроумный механизм, который, несомненно, в сильнейшей степени способствует широкому распространению папоротников.
Описанного выше типа кольца из однорядных клеток, действующего подобно катапульте, еще не было у вымерших папоротниковидных. Его нет также у ряда современных папоротников. Более того, некоторые современные папоротники вообще не имеют никакого кольца и, как уже было сказано выше, оно совершенно отсутствует у семейства ужовниковых.
У тех родов семейства мараттиевых, спорангии которых еще не срослись в синангии (как у ангиоптериса — Angiopteris), имеется примитивное кольцо, напоминающее кольца некоторых палеозойских папоротниковидных. Однако оно не имеет формы кольца, а представляет собой полоску из несколько более крупных клеток с утолщенными стенками, расположенную на конце продольной линии раскрывания спорангия и более или менее поперечно к ней (поперечное кольцо). Конечно, такое кольцо имеет мало общего с типичным кольцом, о механизме действия которого мы рассказали выше. Оно служит для раскрывания спорангия, но эффект катапульты здесь отсутствует. Столь же примитивно кольцо осмундовых, представляющее собой группу толстостенных клеток, расположенную на одной стороне спорангия близ его верхушки.
Если мы теперь обратимся к схизейным, то найдем у них очень хорошо развитое кольцо, заметно отличающееся от подобного образования у остальных современных папоротников.
Оно состоит обычно из одного ряда утолщенных клеток, расположенного непосредственно вокруг верхушки спорангия. Сама верхушка спорангия состоит из одной или нескольких клеток, из чего следует, что кольцо не строго верхушечное. Это, в сущности, поперечное кольцо, расположенное вокруг верхушки. Кольцо способствует продольному раскрыванию спорапгия, но опять-таки без катапультирования. Интересно, что у палеозойских зигоптерисовых кольцо спорангиев было вертикальным, а не поперечным. Из этого можно сделать вывод, что схизейные произошли скорее от их общих предков с зигоптерисовыми, чем непосредственно от зигоптерисовых, и что кольцо у них возникло независимо. Папоротники с аппаратом раскрывания спорангиев типа современных схизейных известны уже с карбонового времени. У адиантовых, которые, по современным представлениям, произошли от предков современных схизейных, кольцо приобрело продольное (вертикальное) положение.
Эволюция кольца спорангия других современных семейств папоротников начинается с косого кольца, которое, следовательно, занимает промежуточное положение между поперечным и продольным (вертикальным). Такое косое кольцо характерно для довольно древних семейств глейхениевых и матониевых. У диптериса мы уже наблюдаем более или менее продольное кольцо, а у полиподиевых кольцо уже вполне продольное и раскрывание спорангия поперечное.
Если, наконец, мы перейдем к третьему большому направлению эволюции современных семейств папоротников, то здесь картина окажется сходной. У циатейных кольцо еще более или менее косое (иногда почти продольное). Косое или иногда почти поперечное кольцо наблюдается также в очень близком семействе гименофилловых. У более подвинутых в эволюционном отношении асплениевых и даваллиевых кольцо приобрело продольное положение и спорангии у них раскрываются поперечной щелью.
Таким образом, эволюция кольца во всех трех ветвях родословного древа папоротниковидных заканчивается наиболее прогрессивным продольным (вертикальным) типом. Преимущество продольпого кольца, вероятно, состоит в том, что один его конец прочно прикреплен к ножке спорангия, которая служит надежной опорой для движений кольца назад и возвращения в исходное положение. Сходство с продольным кольцом некоторых палеозойских папоротников не очень большое, так как у последних они состояли из нескольких (до 8) рядов клеток и, вероятно, функционировали несколько иначе. Продольное кольцо возникало в процессе эволюции независимо несколько раз и в настоящее время является наиболее распространенным типом.
А теперь попытаемся выяснить, в чем именно заключается биологическое значение перехода от поперечного кольца к косому, а этого последнего к продольному. Исследования английского птеридолога (специалиста по папоротникам) Ф. Бауэра показали, что эволюция кольца спорангия была тесно связана с эволюцией всего соруса.
Раскрывание спорангиев с поперечным кольцом происходит в продольном направлении. Продольное раскрывание мы наблюдаем у осмундовых, схизейных и глейхениевых, т. е. у семейств с относительно еще крупными спорангиями, которые или еще не собраны в сорусы, или образуют примитивные сорусы, как у глейхениевых. На этой стадии эволюции продольное раскрывание достаточно эффективно. Но с образованием градатных и особенно смешанных сорусов дело меняется. При большом числе более скученных спорангиев продольное раскрывание становится неудобным по чисто механическим причинам и поэтому сменяется боковым (сначала косым, а затем поперечным).
Косое раскрывание характерно для градатных сорусов. Удлиненное ложе градатных сорусов облегчает боковое (в данном случае косое) разверзание спорангиев, так как здесь все спорангии раскрываются наружу и тем самым отсутствуют физические препятствия для рассеивания спор. Это хорошо видно у циатейных и особенно у гименофилловых. Косое раскрывание спорангиев является в этом случае достаточно эффективным. Однако ситуация резко меняется с переходом градатных сорусов в смешанные. В смешанных сорусах (где более многочисленные мелкие спорангии тесно скучены, а ложе обычно плоское) непрерывное (полное) косое кольцо переходит в более или менее прерванное в месте прикрепления ножки неполное продольное кольцо и раскрывание тем самым становится поперечным, как это хорошо видно, например, у щитовника (Dryopteris). Так как длинная ножка зрелого спорангия приподнимает его над окружающими более молодыми спорангиями, то продольное кольцо может без всяких физических помех полностью закрутиться на обратную сторону и вновь с силой вернуться в исходное положение, разбрасывая таким образом споры. В сорусах промежуточного типа между градатными и смешанными или между простыми и смешанными наблюдается промежуточный тип кольца и промежуточный способ раскрывания спорангия.
В процессе эволюции спорангия и всего соруса и совершенствования механизма его раскрывания число спор в каждом спорангии постепенно уменьшается. Важно подчеркнуть, что речь идет об уменьшении количества спор, производимого не спорофитом в целом, а лишь каждым отдельным спорангием. Число спор, производимых спорофитом, не уменьшается (оно может даже увеличиваться), так как в ходе эволюции равноспоровых папоротников количество спорангиев имеет определенную тенденцию возрастать.
В примитивных, крупных спорапгиях количество спор еще довольно велико. Так, у некоторых ужовниковых насчитывают до 15 000 спор (обычно 1500—2000), а у мараттиевых хотя и меньшее количество спор, но тоже достаточно большое — от 1450 до 7500. Более значительное уменьшение количества спор мы наблюдаем у глейхениевых (1024, 512, 256, 128), осмундовых (от 512 до 128), гименофилловых (512, 256, 128, а иногда 64 и даже 32) и схизейных (256, чаще 128, иногда всего 64). У всех остальных папоротников лишь редко 128 спор (Cheiropleuria), обычно 64 (чаще всего), 48 или 32 споры, редко 16 и еще реже 8 спор. У разноспоровых папоротников число микроспор 64. Из мегаспор полного развития достигает только одна.
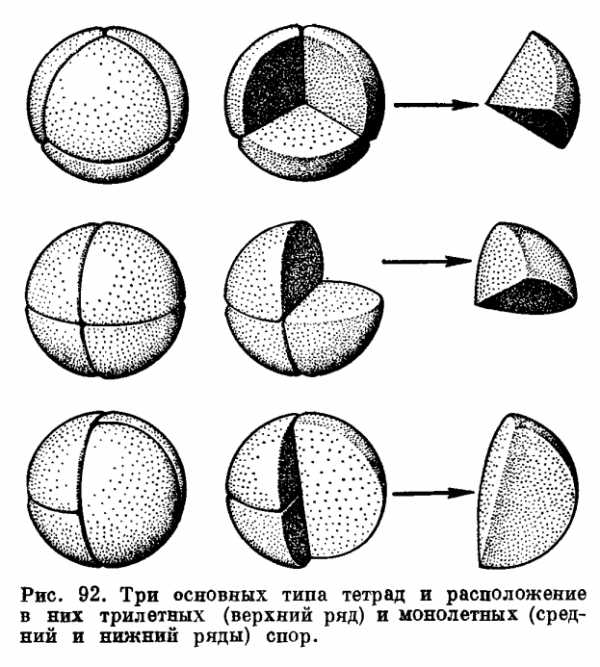
Как мы уже знаем из вводной главы этого тома, в спорах содержится вдвое меньшее число хромосом, чем в клетках спорофита. Мы знаем также, что это уменьшение числа хромосом происходит в процессе деления материнской клетки споры. Из каждой материнской клетки образуются четыре споры, или тетрада спор (рис. 92). На первых стадиях развития эти четыре споры еще тесно связаны между собой своей внутренней стороной, или так называемым проксимальным (от лат. proximus — ближайший) полюсом. Противоположный, обращенный кнаружи полюс называют дистальным (от лат. distalis — дальше отстоящий). Когда тетрада распадается, то каждая отдельная спора на своем проксимальном полюсе несет след своей связи с соседними спорами тетрады — тетрадный рубец, на котором расположена щель. Через эту щель на ранних стадиях развития тетрады осуществляется цитоплазматический обмен между спорами. Затем после расхождения спор щель является единственным местом контакта цитоплазмы споры с внешней средой (ведь спора живет и ей необходимо дыхание) и наконец через щель спора прорастает.
У папоротников имеются два типа тетрадного рубца — трехлучевой и однолучевой. Споры с трехлучевым тетрадным рубцом называют трехрубцовыми или трилетными. Споры с однолучевым тетрадным рубцом называют однорубцовыми или монолетными. Эти два типа спор различаются также по своей геометрической форме. Трилетные споры характеризуются радиальной симметрией и имеют форму более или менее шарообразных тетраэдров, и для простоты их называют тетраэдрическими. Монолетные споры имеют только две плоскости симметрии. Они более или мепее билатеральные — бобовидные или эллипсоидальные. От чего же зависят форма споры и тип ее тетрадного рубца? Как показали специальные исследования (Ф. Бауэр, Н. Маренго, А. Н. Сладков), форма споры и тип ее тетрадного рубца определяются на стадии тетрады. Деление материнской клетки споры происходит разными способами. В случае образования трилетных спор материнская клетка дает начало четырем тетраэдрически симметричным спорам, соприкасающимся одна с другой гранями пирамидальной проксимальной стороны. Каждая такая спора соприкасается с тремя другими спорами, и на ее проксимальном полюсе образуются три радиально расходящихся рубца, составляющие вместе один трехлучевой рубец.
При образовании монолетных спор материнская клетка делится на четыре удлиненные билатерально-симметричные «четверти сферы», которые на стадии тетрады соприкасаются гранями клиновидной проксимальной стороны. Такие билатеральные споры могут образоваться в тетрадах двух типов — изобилатеральных и крестообразных. В изобилатеральных тетрадах длинные оси всех четырех спор расположены параллельно по отношению друг к другу и таким образом каждая спора соприкасается лишь с двумя соседними спорами. В крестообразных тетрадах споры образуют две пары, причем длинная ось одной пары образует прямой угол с длинной осью другой пары. В таких тетрадах каждая спора соприкасается со всеми остальными, причем одной половиной проксимальной стороны с одной спорой, а другой половиной — с двумя. В результате у споры образуется простой однолучевой рубец.
Билатеральные монолетные споры произошли из тетраэдрических трилетных спор, причем это превращение трилетных спор в монолетные произошло независимо в разных линиях эволюции. Монолетные споры появляются уже у некоторых относительно примитивных семейств. Они встречаются у мараттиевых (мараттия — Marattia, данея — Danaea), схизейных (схиаея — Schizaea), некоторых глейхениевых и циатейных, у адиантовых (около половины представителей подсемейства виттариевых — Vittarioideae), а также характерны для древнего рода диптерис. Такие же высокоподвинутые семейства, как многоножковые и асплениевые, характеризуются уже монолетными спорами.
Если монолетные споры произошли из трилетных, то это, очевидно, связано с тем, что в процессе эволюции исходный тетраэдрический тип тетрады дал начало двум другим типам — изобилатеральному и крестообразному. В обоих случаях возникает монолетная спора, но, как показал А. Н. Сладков (1962, 1970) на основе моделирования спор и тетрад, это происходит двумя разными способами. В случае превращения тетраэдрических тетрад ,в изобилатеральные монолетная билатеральная спора возникает путем редукции и исчезновения двух лучей первоначального трехлучевого тетрадного рубца. Когда тетраэдрическая тетрада превращается в крестообразную, монолетная спора образуется за счет редукции и исчезновения одного луча трехлучевой тетрады трилетных спор и увеличения угла между оставшимися лучами до 180°.
В чем заключается биологическое преимущество монолетных спор по сравнению с трилетными? К сожалению, на этот вопрос можно ответить только догадками. В случае монолетных спор деление материнской клетки происходит более простым способом и достигается путем образования меньшего числа перегородок. Возможно, это обстоятельство играло определенную роль в переходе тетраэдрических спор в билатеральные. Но было, вероятно, не менее важным изменение формы споры и укрепление ее рубца. Не лишено вероятности, что рассеивание и прорастание монолетных спор происходит легче, чем трилетных. Однако для выяснения этого вопроса нужны специальные исследования.
Переход трилетного типа в монолетный происходит без большого труда. Это видпо из того, что некоторые папоротники, нормально производящие трилетные споры, могут наряду с последними производить также некоторое количество монолетных спор, что наблюдалось, например, у ужовника (Ophioglossum), гроздовника, ангиоптериса и птериса (Pteris). Но нередко наблюдается и обратное — наличие некоторого количества трилетпых спор у папоротников, нормально образующих только монолетные споры. Это можно наблюдать у мараттии и схизеи вильчатой (Schizaea dichotoma) или у щитовника пильчатого (Dryopteris serrata). Более того, в ряде случаев наблюдаются промежуточные формы между трилетными и монолетными спорами. Так, например, у диптериса китайского (Dipteris chinensis) и птериса критского (Pteris cretica) можно видеть сильное удлинение одного из трех лучей тетрадного рубца при одновременном укорочении, а затем и исчезновении двух других лучей. При наблюдении таких промежуточных спор возникает мысль, что их образование обязано промежуточным типам тетрад. И действительно, по исследованиям А. Н. Сладкова (1962), моделирование промежуточных форм спор и составление из таких моделей тетрад показали, что тетрады также получаются промежуточные, причем двух типов: переходные от тетраэдрических к изобилатеральным и переходные от тетраэдрических к крестообразным.
Из двух основных слоев оболочки спор — интины (эндоспория) и экзины (экзоспория) — наибольший интерес представляет последний. Экзина интересна тем, что, как и у подавляющего большинства других высших растений, она отличается большим разнообразием строения. У разных групп папоротников она различна по своей толщине и, что гораздо важнее, по своей структуре. При исследовании спор папоротников с помощью сканирующего электронного микроскопа споры большинства папоротников обнаруживают поразительное разнообразие экзины. На таблицах 22, 24, 27 и 28 показаны некоторые из наиболее интересных спор. Строение экзины, особенно когда ее изучают с помощью электронного микроскопа, все шире используют для решения вопросов систематики и эволюции папоротников, как ныне живущих, так и вымерших.
Споры большинства папоротников (особенно асплениевых) имеют также дополнительную наружную оболочку, откладываемую на поверхности уже сформировавшейся экзины так называемым плазмодием. Плазмодий (или периплазмодий) образуется в результате разрушения клеток тапетума (выстилающего слоя спорангия). У более примитивных папоротников с многослойными стенками спорангия тапетум образуется из клеток самого внутреннего слоя стенки, а у папоротников с однослойной стенкой он образуется из самой спорогенной ткани. Плазмодий служит питательной средой для развивающихся спор. В него споры погружены во время созревания. Так образуется дополнительный слой оболочки споры — перина (периспорий). Перина обычно резко отличается от экзины, бывает неплотно с ней соединена и более или менее легко разрушается и отпадает. В ископаемом состоянии она не сохраняется. Перина может быть гладкой, сетчатой, шиповатой и т. д. и в некоторых случаях достигает значительной сложности. Поэтому она также может иметь таксономическое значение. Наличие такого рода перины рассматривается как признак специализации. Как показали электронномикроскопические исследования французского палинолога Б. Люгардона (1974), перина имеется не только у асплениевых. Оказалось, что и у остальных папоротников имеется перина, но она образует столь тонкий слой, что не может быть обнаружена с помощью светового микроскопа.
Любопытной особенностью спор некоторых папоротников является наличие в них хлоропластов. У большинства папоротников споры содержат только пропластиды и поэтому они желтые или коричневые (редко почти черные), однако споры осмундовых, гименофилловых и граммитисовых и некоторых асплениевых (оноклея и страусник) содержат значительное число хлоропластов и имеют поэтому зеленый цвет. Биологическое значение хлоропластов в спорах не вполне ясно. Но приуроченность спор с хлоропластами к характерным эпифитам тропических лесов (гименофилловые, граммитисовые) позволяет предполагать, что наличие хлорофилла способствует быстрому прорастанию спор, что, несомненно, облегчает расселение. У обитателей умеренных зон (оноклея, страусник, осмундовые) зеленые споры продуцируются только весной, когда выпадает наибольшее количество дождей. В обоих случаях связь с избыточной влажностью очевидна. Зеленые споры прорастают сразу, без периода покоя.
Наше знакомство со спорами папоротников было бы неполным, если бы мы не рассмотрели вопрос о происхождении разноспоровости. Разноспоровость, т. е. дифференциация спор на микро- и мегаспоры (иногда их называют мужскими и женскими спорами), возникала в процессе эволюции песколько раз. Разноспоровые группы известны как среди вымерших, так и среди ныне живущих папоротников. У одного из современных папоротников, а именно у монотипного северо-восточноавстралийского рода платизома (Platyzoma), составляющего отдельное семейство платизомовых (Platyzomataceae), уже давно была обнаружена зачинающаяся разноспоровость, что полностью подтверждается новейшими исследованиями. У этого папоротника одни спорангии крупнее, а другие мельче. В первых содержится до 16 относительно крупных спор, а во вторых — до 32 мелких спор. Мелкие споры при прорастании дают начало гаметофиту, на котором развиваются только антеридии. Из крупных спор развиваются гаметофиты, на которых в молодости образуются антеридии, а позже только архегонии. Таким образом, если мелкие споры можно назвать микроспорами, то более крупные споры не являются еще настоящими мегаспорами. Разноспоровость у платизомы еще не полная, не вполне установившаяся. Но среди современных папоротников есть три семейства, у которых разноспоровость достигла полного развития. Это — марсилеевые (Marsileaceae), сальвиниевые (Salviniaceae) и азолловые (Azollaceae).
Биологическое преимущество разноспоровости перед равноспоровостью заключается в том, что гаметофит развивается внутри споры (становится эндоспорическим) и развивается за счет тех питательных веществ, которые содержатся в споре, особенно обильно — в мегаспоре. Благодаря быстрому развитию сильно редуцированного паразитического гаметофита процесс оплодотворения происходит рано, гораздо раньше, чем у равноспоровых папоротников. Обильная нища, накопленная в мегаспоре, создает также максимально благоприятные условия для развивающегося зародыша.
Биологические преимущества, достигнутые в результате разноспоровости, вероятно, важны в любых условиях среды, но особенно важны в крайних или меняющихся условиях существования. Марсилеевым, сальвиниевым и азолловым биологические преимущества разноспоровости дают возможность существовать в условиях высыхающих время от времени болот и прудов. Чередующиеся дожди и образование временных водоемов дает им возможность быстро завершить весь репродуктивный цикл.
Таковы основные морфологические и биологические особенности спор. В последние десятилетия всестороннему изучению спор папоротников, как и спор вообще, уделяется все большее внимание. Ведь производство спор есть завершающий этап в развитии спорофита. Все структурные и функциональные изменения спорофита, происходящие в ходе эволюции, связаны с необходимостью размножения посредством спор. «Победителями в борьбе за существование» оказываются те, у кого дости гается наибольшая эффективность в производстве, рассеивании и прорастании спор.
В заключение этого раздела нужно сказать несколько слов о так называемой апоспории — чрезвычайно интересном в биологическом отношении явлении, когда гаметофит образуется не из споры, а из вегетативной клетки или клеток спорофита. Вполне понятно, что такой гаметофит будет иметь одинаковое с клетками спорофита число хромосом. В природе апоспория встречается не часто, но ее обычно бывает нетрудно вызвать искусственно. Легче всего это удается, если отрезанные листья молодого спорофита поместить на агаровую среду. Через некоторое время клетки листа, соприкасающиеся с агаром, дадут начало нитям, которые в дальнейшем превращаются в типичный (но диплоидный) гаметофит.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
Папоротникообразные
Папоротникообразные.
Отдел Папоротнико видные(полифодияфита, птелидофита)
7000 тысяч видов
Происхождение: возникли одновременно с хвощами и плаунами и развивались одновременно с ними. Бурный расцвет в карбоне. Древовидные формы,многочисленные, современно живущие папоротники многообразные,объединяются в большое количество родов.
География: встречаются пркатически везде, наибольшее разнообразие в тропиках. Встречаются в умеренной зоне и даже на севере.
Экология: большинство папоротников произрастаю в лесных сообществах в наземных. Есть луговые представители, в низинах, на скалах. Эпифиты в тропической зоне, эпифиллы. Существуют водные папоротники. Есть земноводные.
Жизненные формы: травы, многолетние, деревья, лианы.
Строение тела: преобладающее поколение спорофит, его строение. Стебель-это корневище( у большинства травянистых папоротников). У тропических форм надземный прямостоячий,одет массой опасных листев.
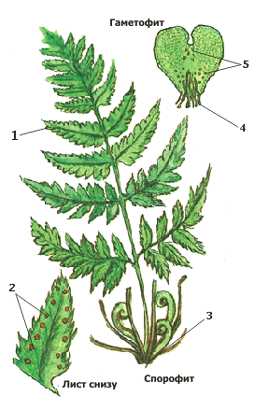
Схема строения папоротника: 1 — Лист, 2 — Сорусы, 3 — Корневище, 4 — Ризоиды, 5 — Антеридии и архегонии

Цикл развития папоротниковидных: 1 — Спорофит (2n), 2 — Споры (n), 3 — Гаметофит (n), 4 — Яйцеклетка (n), 5 — Сперматозоид (n), 6 — Зигота (2n)
Строение стебля сложное и разнообразное. Наблюдается несколько типов: протостелла, сифоностелла, диктиостелла, эостелла.
Корни только придаточные,листья папоротника отличаются от листьев плаунов и хвощей по всем параметрам: форма, размер и происхождение.
Если у хвощевых макрофилия, но листья образовались в результате срастания мелких боковых веточек, то макрофилия у папоротников это результат уплощения( кладофикация) целых больших ветвей. Потому листья современных папоротников имеют черты побегов. Они растут верхушкой, долго, разиваются медленно, 2 года в почке и появляется лист улитка.
Листья имеют разнообразные формы и размеры и виды рассечения. У тропических форм древовидных видов нескольких метров длинной, многократно рассеченные, а у папоротника рода азолла. Самое древнее ветвление дихотомическое, перистое, сетчатое. Основная функция листа фотосинтез. Основная функция спороношение . Спорангии располагаются либо по краям листа на верхушке с нижней стороны.
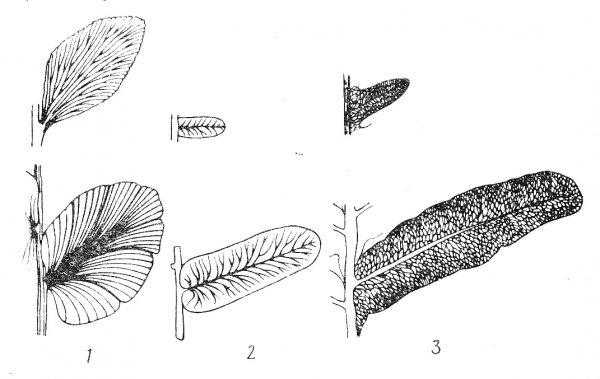
1-веерообразное; 2-перистое; 3-сетчатое.
Спорангии могут срастаться в синангии или группируются в соруса.Строение соруса.
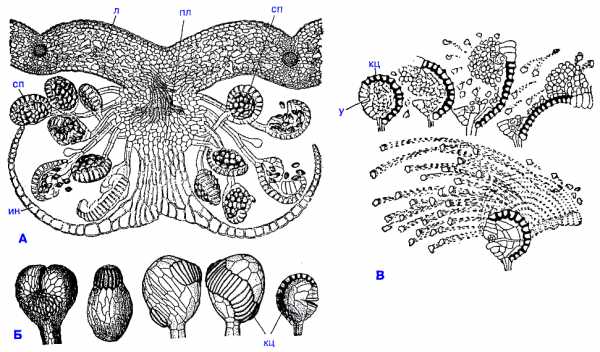
Строение сорусов и спорангиев папоротников:
А - поперечный разрез соруса щитовника мужского (Dryopteris filix-mas) ;
Б - разнообразие спорангиев;
В - раскрывание спорангиев; л - лист; пл - плацента; сп - спорангий; кц - кольцо; у - устье; ин - индузий.
У некоторых папоротников деморфизм листьев, одни выполняют функцию фотосинтеза, а другие спороношения.
У многих эпифитных папоротников наблюдается деморфизм фотосинтезирующих листьев.
У современных папоротников есть равно и разноспоровые(савиниевые, азоловые) представители.
Равноспоровые папоротники – подавляющее большинство, их гаметофит обоеполый, у примитивных видов он подземный, червеобразный, входит в симбиоз с грибами. Половозрелый развивается медленно. У более продвинутых папоротников гаметофит надземный, зеленый, быстро созревающий, имеет вид пластины иногда сердцевидной формы.
Жизненный цикл.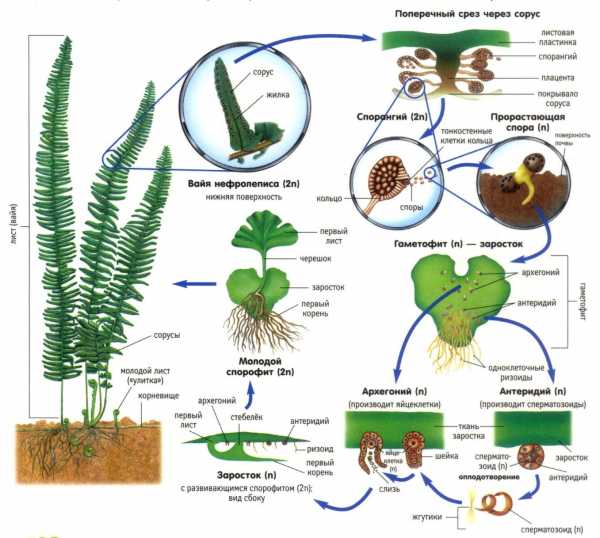
Гаметофиты разноспоровых папоротников раздельнополые, отличаются сильной редукцией( особенно мужской) и развитием мега и микроспорангия.(Сальвиния).
В основании листьев образуются сорусы. Таким образом разноспоровость возникала неоднократно.
Среди ныне живущих выделяют следующие систематические категории.
Ужовниковые.
Маратиевые(тропические декоративные виды)
Полиподиопсиевые(разнообразные): осмундовые, адиантовые, подчешуйниковые, щитовниковые, оноплиевые, сальвиниевые, азоловые.
Значение:
1.Входят в состав растительных сообществ доминируя в нижних ярусах.
Для человека:декоративные, оранжерейные, аквариумные растения, стебли и листья древовидных используются для поделок, крахмал листьев и стеблей используется в пищу. В россии кушают папоротник орляк, в США страусник. Лекарственные: щитовник мужской.
Азола
Удваивает свою биомассу вдвое. Один из самых быстрорастущих растений в мире. В китае азолу используют на рисовых полях. Объект хороший для космонавтов, не требует особых условий, вкусная и полезная.
Азолу можно использовать для отчистки сточных вод.
На нижней стороне папоротника распологаются сорусы каково их значение
Как известно, папоротники являются одной из групп споровых растений. У большинства их видов споры образуются на нижней стороне зелёных листовых пластин внутри специальных вместилищ, которые и называются соросами.
Строение папоротника
Папоротники отличаются по своему строению как от семенных растений, так и от остальных споровых, в число которых входят:
- мхи;
- плауны;
- хвощи.
Основным отличием является строение их веток и листьев. Если у других групп сосудистых растений ветки и листья размещаются в разных плоскостях и фактически отделимы друг от друга, то у папоротников они располагаются в одной плоскости. От других споровых растений папоротники отличаются также тем, что органы размножения (т.е. споры) появляются у них не на верхней части стебля, а под листьями.
Споры собираются в соросах, где дожидаются своего созревания. После созревания они становятся тёмно-коричневого, а иногда и черного цвета. Затем споры падают в грунт и начинают прорастать.
Циклы роста папоротника
После попадания в грунт, споры папоротников активизируются и идут в рост. Однако из них не сразу прорастают те формы папоротников, которые мы привыкли видеть.
В первой фазе папоротники имеют очень маленький размер, всего лишь несколько миллиметров, и при этом они разнополые. Спустя некоторое время, внутри них начинают развиваться специальные клетки, которые называют гаметами. Именно поэтому на первой стадии папоротник называют гаметофитом.
Когда мужские и женские гаметы соединяются, из них вырастают уже взрослые особи папоротников. Это второе состояние называется спорофитом. Именно под листьями спорофитов мы и находим сорусы.
На территории России встречается около двух десятков видов папоротников, которые, как и в большинстве других регионов умеренной полосы, к зиме сбрасывают свои листья. Однако их подземные корни, которые иногда формируют клубни, при этом остаются живыми. Поскольку папоротники являются многолетними растениями, весной они прорастают заново, и цикл их размножения и развития возобновляется.
Споры и спорангии папоротника
Содержание статьи
Спорангии папоротника
Папоротники имеют самое сложное строение по сравнению с остальными споровыми. Их развиваются под землей. Они называются корневищами. На поверхности почвы вырастают листья. Сначала они спирально закручены. По мере роста они могут достигать значительных размеров. Так выглядит растение бесполого поколения.
Что такое спорангии папоротников, и где они находятся? Эти структуры хорошо заметны на нижней стороне их листьев. Спорангии имеют вид небольших коричневых бугорков, прикрытых особыми чешуйками. Они называются «покрывальцами». После созревания споры попадают в почву и прорастают. В результате формируется особь полового поколения — заросток. Это пластина сердцевидной формы зеленого цвета, диаметр которой достигает лишь сантиметра. Подобно хвощам и плаунам, заросток папоротника развивается отдельно от спорофита. Он растет на поверхности влажной почвы, прикрепляясь к ней при помощи ризоидов.
Развитие гаметофита
Споры папоротника развиваются только во влажной почве. Их распространение происходит при помощи воды, ветра, насекомых и птиц. В результате прорастания споры формируется пластинка. В поперечном сечении ее размер едва достигает 3 мм. Заросток папоротника прикрепляется к земле с помощью ризоидов, что обеспечивает еще и дополнительное питание растения.
На нижней стороне этой структуры располагаются половые органы — гаметангии. Эти структуры также имеют микроскопические размеры, которые даже сложно представить. Они бывают двух типов. Мужские называются антеридии. В них созревают сперматозоиды. Это половые клетки, которые имеют жгутики. Но передвижение сперматозоидов возможно только при наличии воды. Поэтому наличие влаги является обязательным условием для осуществления процесса оплодотворения у всех споровых растений. Женские половые органы называются архегонии. В них формируются неподвижные яйцеклетки.
Для папоротников характерно перекрестное оплодотворение. Это объясняется тем, что созревание антеридиев и архегониев происходит в разное время. Поэтому слияние гамет обычно происходит между разными проростками.
Размножение грибов
Бесполое размножение грибов происходит благодаря особым спорам, которые развиваются на специальных ветвях мицелия. Споры могут быть экзогенными и эндогенными. Что такое спорангии грибов? Это особые клетки, которые также называются спорангиоспорами. Экзогенные споры образуются в органах, которые называются конидиеносцами, а споры – конидиями. Высшие грибы размножаются только конидиями, низшие чаще всего спорами.
Спорангии образуются на спорангиеносцах. Это особые ветви грибницы, которые отличаются ограниченным приростом, большей толщиной и другими особенностями.
Кстати, плесневые грибки выделяют токсические вещества, называемые микотоксинами, которые представляют собой сильнодействующий яд, способный причинить вред человеку и животным. Для этого совсем не обязательно скушать заплесневелый хлеб или другую пищу. Достаточно развернуть пакет или открыть судочек с заплесневевшей едой и случайно вдохнуть споры с воздухом или прикоснуться к плесени.
Надеемся, теперь вы понимаете, что такое спорангии, и знаете, что в них образуются споры, с помощью которых растения размножаются. Иногда споры могут быть ядовитыми.
Размножение
Спорангии находятся на нижней стороне листа, собраны кучками (сорусы). Сверху сорусы прикрыты покрывальцем (кольцо). Споры рассеиваются при разрыве стенки спорангия, а кольцо, оторвавшись от тонкостенных клеток, ведёт себя подобно пружине. Число спор на одном растении достигает десятков, сотен миллионов, иногда миллиардов.
Лист папоротника с нижней стороны
На влажной почве споры прорастают в маленькую зелёную сердцеобразную пластинку величиной несколько миллиметров. Это заросток (гаметофит). Он располагается почти горизонтально к поверхности земли, прикрепляясь к ней ризоидами. Заросток обоеполый. На нижней стороне заростка образуются женские и мужские половые органы (мужские — антеридии, женские — архегонии).
Оплодотворение происходит в водной среде (во время росы, дождя или под водой).
Мужские гаметы — сперматозоиды подплывают к яйцеклеткам, проникают внутрь и гаметы сливаются.
Происходит оплодотворение, в результате чего образуется зигота (оплодотворённая яйцеклетка).
Из оплодотворённой яйцеклетки формируется зародыш спорофита, состоящий из гаустории — ножки, которой он врастает в ткани заростка и потребляет из него питательные вещества, зародышевого корешка, почки, первого листа зародыша — «семядоли».
Со временем из заростка развивается растение папоротник.
Схема развития папоротника
Таким образом, гаметофит папоротников существует независимо от спорофита и приспособлен к обитанию в условиях увлажнения.
Спорофитом является всё растение, которое вырастает из зиготы — типичное сухопутное растение.
biouroki.ru
Общие понятия
Спорангии — это многоклеточные (у высших растений) и одноклеточные (у водорослей) органы, в которых образуются споры. Вы когда-нибудь видели плесень на хлебе? В ней можно различить малюсенькие черные точечки, которые тоже являются спорангиями. В одном таком спорангии может содержаться до 50 тысяч спор, каждая из которых воспроизводит до сотни миллионов новых спор за несколько дней! Именно поэтому плесень размножается невероятно быстро.
Споры в спорангиях похожи на мелкие шарики, покрытые оболочкой. Спорангии хвощевидных, плауновидных, папоротниковидных развиваются на спорофиллах и могут быть собраны в сорусы (группы) или быть одиночными.
Папоротники
ПАПОРОТНИКИ (папоротниковидные), отдел высших споровых растений. Имеют древнее происхождение. Возникли в девоне; в карбоне древовидныепапоротники вместе с ископаемыми плаунами и хвощами господствовали в растительном покрове Земли. Ныне насчитывается ок. 300 родов и более 10 тыс. видов папоротников. Они играют меньшую роль в формировании растительного покрова, однако широко распространены по всему земному шару и встречаются в самых различных местообитаниях. Наибольшее их разнообразие наблюдается в тропиках и субтропиках, где встречаются папоротники травянистые, древовидные, лиановые, водные, папоротники-эпифиты и др. Папоротники умеренных широт – многолетние травянистые растения, произрастающие во влажных тенистых лесах, оврагах, на болотистых лугах, берегах водоёмов, в трещинах скал. Для папоротников характерно чередование поколений. Доминирует бесполое поколение, или спорофит.
По размерам папоротники варьируют от крупных тропических древовидных форм (выс. до 25 м, диам. ствола до 50 см) до крошечных растений длиной в несколько мм. Стебли папоротников различаются по форме и анатомическому строению. Прямостоячие стволы древовидных папоротников у основания обычно снабжены многочисленными воздушными корнями. придающими им устойчивость. Стебли травянистых папоротников, особенно молодые, покрыты волосками или чешуйками. Листья папоротников, которые часто называют вайями, весьма разнообразны. Их длина колеблется от нескольких миллиметров до 30 м. Обычно лист имеет черешок и пластинку разной формы (простую, перистую, дважды или многократно перистую и т. д.). Листья, как правило, совмещают две функции – фотосинтеза и спороношения, лишь у немногих видов они дифференцированы на фотосинтезирующие (стерильные) и несущие спорангии (фертильные). На определённой стадии развития спорофита начинается спороношение (в биологическом смысле оно аналогично цветению). Спорангии расположены, как правило, на нижней стороне листа или по его краю, у большинства папоротников они сгруппированы в компактные кучки – сорусы. Часто сорусы прикрыты защитными органами – покрывальцами (индузиями). Спорангии одного соруса у разных видов созревают или почти одновременно, или последовательно. При созревании спорангии вскрываются и споры высыпаются. Из множества спор, производимых взрослым папоротником, лишь небольшая их часть попадает в благоприятные условия, прорастает, давая начало половому поколению – гаметофиту, или заростку. Для прорастания спор большинства папоротников, кроме влажности и тепла, нужен свет. У разноспоровых папоротников гаметофит однополый, у равноспоровых – обоеполый. Многие папоротники очень декоративны и используются для украшения жилищ и в оранжереях. В Юго-Восточной Азии молодые проростки широко используют в пищу. Корневища папоротника мужского служат источником лекарственного сырья.
sbio.info
Щитовник мужской — Википедия
| Щитовник мужской | |||
|---|---|---|---|
| Общий вид растения, Польша | |||
| промежуточные ранги
| |||
| Dryopteris filix-mas (L.) Schott (1834) | |||
Щито́вник мужско́й, или Па́поротник мужской (лат. Dryópteris fílix-mas) — один из самых распространённых папоротников вообще и самый распространённый папоротник умеренной климатической зоны земного шара; вид рода Щитовник (Dryopteris) семейства Щитовниковые (Dryopteridaceae).
Широко распространённое предание или суеверие о цветке (огненном цветке) папоротника, который нужно найти в ночь на Ивана Купала, связано как раз со щитовником мужским. Кто в эту ночь находил такой цветок, тому открывались подземные клады, дар предвидения. «Огненный» цветок якобы мог сделать любого человека невидимым, дать власть над тёмными силами, сделать его сказочно богатым или счастливым.
Щитовник мужской относится к роду Щитовник (Dryopteris Adans.) — из семейства (или, у некоторых авторов, подсемейства) Щитовниковые (Dryopteridaceae) и является одним из самых широко распространённых видов этого рода как в природе, так и в искусственном озеленении.
Родовое название Dryopteris буквально переводится как папоротник дубовых лесов (происходит от слов греч. δρυας — «дуб» и πτηρων — «крыло птицы»; некоторые европейские виды этого рода действительно встречаются в дубравах).
Видовой эпитет — filix-mas — дословно значит «папоротник мужской» (от лат. filix — «папоротник» и mas — «мужской»). Это название имеет древнее ритуальное происхождение (римское) и носит сравнительный характер, поскольку наряду с мужским папоротником в лесах встречался (и встречается по сей день) также и папоротник женский, отличающийся значительно более нежными, гнущимися и более мелко перистыми листьями, чем мужской.
Щитовник мужской — один из самых красивых и широко известных лесных папоротников. Он имеет характерного вида короткое и толстое, косо поднимающееся вверх корневище, покрытое широкими мягкими чешуями (коричневыми или чёрными) и остатками листовых черешков.У щитовника мужского вильчатое жилкование .
На вершине корневища располагается розетка крупных листьев с дважды перистой пластинкой. Длинные (ланцетной формы) прямостоячие двоякоперисторассечённые вайи собраны в воронковидный пучок. Листовые черешки короткие, толстые, густо усаженные короткими бурыми чешуйками. Если повернуть лист нижней стороной к себе, можно увидеть на сегментах листовой пластинки по пять — восемь сорусов, расположенных точно в местах разветвления питающих жилок и прикрытых сверху почковидными покрывальцами. Сорусы расположены в два ряда. Споры (видимые только под мощным микроскопом) имеют почковидную форму с гребешочками и усечёнными бородавочками по всей поверхности.
Листья щитовника мужского очень декоративны. Они могут достигать длины одного или даже полутора метров, однако растут медленно, долго не грубеют и могут быть легко повреждены. Появляясь в виде листовых зачатков в форме узкого кольца вокруг точки роста на вершине корневища, они перезимовывают в таком виде первый раз — и только на второе лето принимают характерную для всех папоротников улиткообразную форму. Молодые листья появляются в самом центре розетки и таким образом максимально защищены от любых внешних воздействий. Густое покрытие чешуйками и свёрнутое улиткообразное положение молодых листьев лучше всего предохраняют нежные ткани растущей верхушки листа от высыхания и любых механических повреждений. В таком улиткообразном состоянии пучок молодых листьев проводит ещё один год. И только весной третьего года молодые листья быстро развёртываются в густую розетку и достигают своего полного развития. Таким образом, в разгар периода вегетации на каждом растении одновременно можно найти все три формы листьев трёхгодового цикла: это годовалые зачатки, двухгодовалые улитки и полностью развитые прямостоячие зрелые вайи. Сформированные листья живут один сезон, выполняют вегетативную функцию, а также функцию размножения — и той же осенью увядают. Но к тому моменту уже вызревают и рассеиваются споры, из которых в благоприятных условиях той же осенью вырастает и уходит под снег на зимовку гаметофит (примитивный обоеполый заросток нового растения) сердцевидной формы, обильно покрытый железистыми волосками.
Отличается от кочедыжника женского (Athyrium filix-femina) формой сорусов (у женского папоротника они продолговатые), от других видов щитовника — характером края листа, окраской плёнок на листовом черешке.
Спороносит с середины до конца лета. Средний возраст спороносящих экземпляров свыше 30 лет[1].
Все части растения содержат фенольные соединения и антоцианы. Корневища, кроме того, — тритерпеноиды, витамины группы B, дубильные вещества (7—8 %), высшие алифатические спирты и высшие жирные кислоты. В листьях найдены эфирное масло (0,144 %), витамин C, флавоноиды, высшие жирные кислоты, в том числе линолевая, пальмитиновая, олеиновая, линоленовая, стеариновая; липиды[2].
 Заросли щитовника весной
Заросли щитовника весной Щитовник мужской можно в полной мере назвать космополитом среди растений. Он освоил самые обширные территории произрастания — его можно встретить от Гренландии и Скандинавии до Мексики и Средиземноморья. На территории России его ареал не менее широкий: его можно найти от Кольского полуострова на севере европейской части до горно-лесных районов Кавказа, Предуралья, Урала и юга Сибири. Встречается Щитовник мужской также и в Средней Азии. В горах он поселяется в буковых, еловых, пихтовых и арчовых лесах, поднимаясь местами до альпийского и горно-тундрового пояса. Может расти даже в Арктике — по защищённым от ветра и прогреваемым летом южным склонам, засыпаемым зимой мощным снежным покровом. Однако, основная часть его ареала находится именно в лесной зоне, где он встречается в хвойных, смешанных и широколиственных лесах.
В лесах умеренной климатической зоны (например, средней полосы России или Северо-Запада) на влажных слабокислых почвах в травяном покрове еловых, пихтовых и широколиственных лесов щитовник мужской часто образует плотные сообщества, вытесняя с благоприятной территории почти все остальные растения. Однако, и одиночные экземпляры тоже встречаются не редко. Излюбленное место заселения щитовника мужского — умеренно заболоченные (или даже полусухие) пожарища, зоны отчуждения железных дорог, просеки, где в условиях пониженной конкуренции можно встретить особенно обширные заросли этого растения.
Однако, несмотря на широкое и почти повсеместное распространение, щитовник мужской внесён в региональные Красные книги России (например, Мурманской области[3] и Бурятии) и Украины. В Донбассе встречается в лесах Славянского и Краснолиманского районов Донецкой, а также в поймах Донца в Луганской областях. В Бурятии вид обнаружен на южном побережье Байкала в дельтах рек Снежной, Выдринной, Переемной и в окрестностях станции Выдрино. Восточнее побережья озера Байкал становится ещё более редким, почти не встречается[4]. Сокращение популяции связано прежде всего с уязвимостью уже образовавшихся сообществ растений, которые плохо переносят вырубку лесов и любое вторжение человека в свою среду. Но не только сообщество в целом, но и каждое растение в отдельности имеет достаточно чувствительные корневища, которые легко повреждаются, но очень трудно восстанавливаются.
Хозяйственное значение и применение[править | править код]
Щитовник мужской является постоянным предметом сбора — как лекарственное сырьё, декоративное садовое растение и компонент для выращивания оранжерейных эпифитов (составная часть эпифитного субстрата, так называемые папоротниковые корни).
Ещё в середине XX века корневища щитовника широко использовали для дубления кож и окраски их в жёлтый цвет[2]. Теперь, правда, только редкие кустари-кожевенники, придерживающиеся старых натуральных рецептур, используют папоротник в этих целях.
Экстракт из осенних листьев пригоден для стимуляции размножения дрожжей при их производстве[2].
Корневища ядовиты для свиней и овец при поедании в большом количестве[2].
Лекарственное применение[править | править код]
Щитовник мужской широко и очень давно известен в качестве лекарственного растения. Лекарственные свойства этого растения были известны уже врачам античной эпохи и Средневековья. В частности, о нём неоднократно упоминается в трудах Диоскорида и Плиния.
Растение ядовито, (в особенности корневище), не исключается возможность отравления! Корневище обладает специфическим запахом и сладковато-терпким вкусом.
В качестве лекарственного сырья используют корневище (лат. Rhizoma Filicis maris), которое выкапывают осенью, отряхивают землю, очищают от корней и листьев и высушивают в тени, в хорошо проветриваемых помещениях или в сушилках при температуре не выше 40 °C[5]. Урожайность корневищ до 5 ц/га[1].
В корневищах содержится 7—8 % дубильных веществ, производные флороглюцина, флавоноиды и эфирное масло. Главными действующими веществами корневища папоротника мужского являются фенольные соединения, состоящие из флороглюцидов различной степени сложности, — мономерные, димерные и тримерные производные флороглюцина[5]: фильмарон (аспидинофиллин), филицин, флаваспидиновая кислота, аспидинол и другие, обладающие выраженным противоглистным действием. Эти вещества в экстракте корневищ вызывают паралич мускулатуры ленточных глистов и червей, которых затем выводили из организма при помощи слабительного. Этот способ противоглистной терапии на данный момент считается устаревшим. В европейской фармакопее этот препарат главным образом применяется в ветеринарии. В фармацевтической промышленности лекарственное сырьё используется для приготовления препаратов от червей-паразитов. В качестве глистогонного средства щитовник мужской официально входил в Государственную фармакопею СССР. Из свежесобранных корневищ получали препарат филиксан.
Однако яды, содержащиеся в корневищах папоротника мужского, токсичны не только для гладкой мускулатуры паразитических червей, но и для центральной нервной системы и сердца человека. При введении в кровь подопытных теплокровных животных (или после всасывания в кровь из кишечника) пороговой токсической дозы вскоре начинаются судороги, а впоследствии и паралич центральной нервной системы и сердца. Именно в силу этих токсических свойств препараты щитовника мужского противопоказаны при заболеваниях сердца, болезнях печени и почек, язвенной болезни желудка и двенадцатиперстной кишки, а также при беременности, истощении и малокровии. Кроме того, при употреблении препаратов папоротника мужского (спустя полчаса после приёма) необходимо давать только солевое слабительное (например, английскую соль), но не касторку, которая резко усиливает всасывание в зоне кишечника и таким образом может спровоцировать сильное отравление.
Использование щитовника в народной медицине значительно шире, чем в официальной. Однако, необходимо помнить, что корневище этого папоротника ядовито и при самолечении может вызвать острое отравление, выражающееся в судорогах, помрачении сознания, усиленном сердцебиении и рвоте.
Споры в традиционной китайской медицине применяют при гематурии и болезнях мочеполовых органов[2].
Культурное растение[править | править код]
Щитовник мужской широко используется в качестве высокодекоративного культурного растения в городском озеленении, на приусадебных участках и в садово-парковом хозяйстве. В Европе известен в культуре во всяком случае с XVII века и имеет не менее двух десятков культурных селекционных форм, сильно отличающихся от природного вида. Однако, косвенным образом можно судить также и о том, что и в Древней Греции, и в Римской империи имело место его употребление в качестве культурного или по крайней мере окультуренного дикорастущего растения. Само по себе слово felix или filix в живой латыни времён императоров имело одно значение, однако очень широко употребляемое как в прямом, так и в переносном смысле слова. Слово filix означало папоротник, или (шире) сорняк, а в переносном смысле — назойливый визитёр, ничтожество, а также нежелательные волосы на теле. Именно второе значение этого слова (сорняк) и позволяет судить о широком распространении этого красивого папоротника с правильной кроной листьев, а также о возможности его декоративного использования в городах и на тех местах в саду, возле каменной ограды или у дома, где его присутствие не только не мешало другим растениям (сорняк можно было не удалять), но и было желательно.
В настоящее время садоводами и озеленителями активно используется более трёх десятков сортов щитовника мужского, отличающихся главным образом размером, формой и плотностью листьев. Некоторые наиболее известные культурные формы:
 Щитовник мужской сорт 'Cristata'
Щитовник мужской сорт 'Cristata' - 'Barnesii' — растения по размеру близкие к природному виду, хотя и кажутся более компактными, собранными, высота до 80—90 см. Листья более узкие и жёсткие, прямые и хорошо держат форму, что создаёт впечатление как бы геометрического растения.
- 'Crispa' — растения с более короткими листьями. Доли каждого листа более широкие, чем у исходного вида и почти соприкасаются друг с другом, образуя как бы цельную листовую пластину. Высота растения не превышает полметра.
- 'Crispa Capitata' — растение компактное, с довольно тонкими и длинными листьями. Отдельные сегменты сильно курчавые и обильно ветвящиеся как в форме вилочек, так и в форме гребней. Растение издалека напоминает крупный пучок петрушки.
- 'Crispa Cristata' — такое же компактное растение, однако каждый сегмент и лист в целом оканчивается завитком, гребешком или неполным веером, что создаёт впечатление курчавости.
- 'Cristata' — крупные растения с более широкими, чем у основного вида листьями. Отличается также и формой окончания каждого пера сложного листа. Как следует из названия сорта, каждый листочек заканчивается не «остриём», а гребешком или веером, от основания постепенно сужаясь и в конце резко разворачиваясь.
- 'Furcata' — растения среднего размера, отдельные доли листа многократно раздваиваются, особенно ближе к кончикам. Сорт, по-видимому, производный от предыдущего.
- 'Grandiceps' — растения крупные, по размеру даже превосходят природный вид. Главный черешок листа на конце обильно ветвится, в результате чего лист напоминает форму гриба со шляпкой или бадминтонную ракетку. Каждое перо сложного листа, изящно выгибаясь и поникая, так же образует веер — или «кристату», как у предыдущего сорта. Выносливый, очень декоративный и неприхотливый сорт, широко распространённый в культуре.
- 'Linearis Congesta' — (форма «компресса») как бы сжатое растение с листьями в три раза короче, чем у природного вида (30—40 см). Отдельные перья листа посажены очень густо, отчего создают впечатление курчавости.
- 'Linearis Polydactyla' — похож на предыдущий сорт. Компактное растение, отдельные перья листа узкие, почти линейные, на конце резко разветвляются, образуя нечто вроде раскрытой ладони руки.
- 'Multicristata' — сорт, в целом близкий к сорту Crispa Cristata, более рослый и широкий. Конец листа в целом и каждого сегмента в отдельности заканчивается правильной формы гребнем. Растение густое, высота куста до 70 см.
- 'Ramo Cristata' — очень сильно веерная форма. И весь лист, и каждое перо в отдельности веерно разветвляется почти от самого основания. Растение в целом совершенно не похоже на исходный вид.
- 'Squamulosa' — компактное, низкое и очень густое растение, доли листа соприкасаются или частично перекрывают друг друга, кроме того, каждая пластинка сильно гофрирована, как если бы лист долго мяли в руках. Высота куста не более 40 см.
Щитовник мужской в природе служит пищей для большого количества диких животных. Особенно листья этого папоротника любит лось. В середине 1970-х годов зоологи Саянского заповедника попытались выяснить, какова роль щитовника в питании крупных травоядных животных. Результаты расчётов по времени и скорости передвижения лосей по лесу оказались удивительными. Обыкновенно эти животные едят практически не останавливаясь и срывая листья деревьев и кустарников на ходу. Средняя скорость передвижения лося по лесу составляет примерно 2,5 метра в минуту. И на пойменных лугах с богатой растительностью, и даже в воде эта скорость практически не менялась. А вот в зарослях щитовника, нежные листья которого, казалось бы, сорвать легче всего, лось буквально «вязнет», почти останавливается и начинает пастись со скоростью всего один метр в минуту, довольно серьёзно прорежая густые заросли папоротника. Это одновременно даёт хороший результат для молодых сеянцев сибирского кедра, которые под сплошным покровом щитовника часто погибают от недостатка света и избыточной влажности.
Патогенные грибы[править | править код]
На щитовнике мужском паразитируют несколько видов аскомицетов из рода Тафрина (Taphrina). В Северной и Центральной Европе, на Британских островах и на Дальнем Востоке встречается Taphrina vestergrenii, в северных и горных регионах Европы — Taphrina athyrii. Оба вида вызывают пятнистость листочков[6].
Таксономическая схема[править | править код]
Синонимы[править | править код]
По данным The Plant List на 2010 год, в синонимику вида входят[7]:
- Aspidium depastum Schkuhr
- Aspidium erosum Schkuhr
- Aspidium expansum D.Dietr.
- Aspidium filix-mas (L.) Sw.
- Aspidium mildeanum Göpp.
- Aspidium nemorale (Salisb.) Gray
- Aspidium opizii Wierzb.
- Aspidium umbilicatum (Poir.) Desv.
- Aspidium veselskii Hazsl. ex Domin
- Aspidium × bohemica Domin
- Aspidium patagonica Diem
- Lastrea filix-mas (L.) C.Presl
- Nephrodium crenatum Stokes
- Nephrodium filix-mas (L.) Rich.
- Polypodium filix-mas L.
- Polypodium heleopteris Borkh.
- Polypodium nemorale Salisb.
- Polypodium umbilicatum Poir.
- Polystichum filix-mas (L.) Roth
- Polystichum polysorum Tod.
- ↑ 1 2 Губанов И. А. и др. Дикорастущие полезные растения СССР / отв. ред. Т. А. Работнов. — М.: Мысль, 1976. — С. 29—30. — 360 с. — (Справочники-определители географа и путешественника).
- ↑ 1 2 3 4 5 Часть I — Семейства Lycopodiaceae — Ephedraceae, часть II — Дополнения к 1—7-му томам // Растительные ресурсы России и сопредельных государств / отв. ред. А. Л. Буданцев; Бот. ин-т им. В. Л. Комарова РАН. — СПб.: Мир и семья-95, 1996. — С. 29—31. — 571 с. — ISBN 5-90016-25-5.
- ↑ Щитовник мужской. Флора Мурманской области (Проверено 21 ноября 2013).
- ↑ Красная книга Бурятии — Щитовник мужской (недоступная ссылка) (Проверено 21 ноября 2013).
- ↑ 1 2 Блинова К. Ф. и др. Ботанико-фармакогностический словарь : Справ. пособие / Под ред. К. Ф. Блиновой, Г. П. Яковлева. — М.: Высш. шк., 1990. — С. 261. — ISBN 5-06-000085-0.
- ↑ Каратыгин И. В. Порядки Тафриновые, Протомициевые, Экзобазидиевые, Микростромациевые. — СПб.: Наука, 2002. — С. 20, 23. — (Определитель грибов России). — ISBN 5-02-026184-X.
- ↑ Dryopteris filix-mas (L.) Schott is an accepted name (англ.). The Plant List (2010). Version 1. Published on the Internet; http://www.theplantlist.org/. Royal Botanic Gardens, Kew and Missouri Botanical Garden (2010). Дата обращения 21 ноября 2013.
- Атлас лекарственных растений. — Веда, 1981.
- Гладкова В. Н. Подсемейство щитовниковые (Dryopteridoideae) // Жизнь растений: в 6 тт. — Т. 4. Мхи. Плауны. Хвощи. Папоротники. Голосеменные растения. / Под ред. И. В. Грушвицкого и С. Г. Жилина. — М.: Просвещение, 1978. — С. 227—229.
- Губанов И. А. и др. 9. Dryopteris filix-mas (L.) Schott — Щитовник мужской // Иллюстрированный определитель растений Средней России. В 3 т. — М.: Т-во науч. изд. КМК, Ин-т технолог. иссл., 2002. — Т. 1. Папоротники, хвощи, плауны, голосеменные, покрытосеменные (однодольные). — С. 83. — ISBN 8-87317-091-6.
- Коновалова Т. Ю. и др. Папоротники для сада. — М.: Кладезь, 2004.
- Меньшикова З. А. и др. Лекарственные растения в каждом доме. — М.: Адонис, 1993.
- Смирнов А. Мир растений — М.: Молодая гвардия, 1982.
- Gardeners’ Encyclopedia of Plants & Flowers. — Dorling Kindersley Limited, London, 1995.
Папоротниковидные — Википедия
Папоротникови́дные, или па́поротники, (лат. Polypodióphyta) — отдел сосудистых растений, в который входят как современные папоротники, так и одни из древнейших высших растений, появившихся около 405 млн лет назад в девонском периоде палеозойской эры. Гигантские растения из группы древовидных папоротников во многом определяли облик планеты в конце палеозойской — начале мезозойской эры.
Современные папоротники — одни из немногих древнейших растений, сохранивших значительное разнообразие, сопоставимое с тем, что было в прошлом. Папоротники сильно различаются по размерам, жизненным формам, жизненным циклам, особенностям строения и другим особенностям. Внешний облик их настолько характерен, что люди обычно называют всех их одинаково — «папоротники», не подозревая, что это самая большая группа споровых растений: существует 48 семейств, 587 родов и 10 620 видов папоротниковидных[1]. Разнообразие форм листьев, удивительная экологическая пластичность, устойчивость к переувлажнению, громадное количество производимых спор обусловили широкое распространение папоротников по земному шару[2]. Папоротники встречаются в лесах — в нижнем и верхнем ярусах, на ветвях и стволах крупных деревьев — как эпифиты, в расщелинах скал, на болотах, в реках и озёрах, на стенах городских домов, на сельскохозяйственных землях как сорняки, по обочинам дорог. Папоротники — вездесущи, хотя и не всегда привлекают внимание. Самое большое их разнообразие — там, где тепло и сыро: тропики и субтропики.
У папоротников ещё нет настоящих листьев, но они сделали в их направлении первые шаги. То, что у папоротника напоминает лист — вовсе не лист, а по своей природе — целая система ветвей, да ещё расположенных в одной плоскости. Так это и называется — плосковетка, или вайя, или, ещё одно название, — предпобег. Несмотря на отсутствие листа, у папоротников есть листовая пластинка. Этот парадокс объясняется просто: их плосковетки, предпобеги претерпели уплощение, в результате которого появилась пластинка будущего листа — почти не отличимая от такой же пластинки настоящего листа, однако папоротники эволюционно ещё не успели разделить свои вайи на стебель и лист. Глядя на вайю, трудно понять, где — на каком уровне ветвления — заканчивается «стебель», и где начинается «лист», но листовая пластинка уже есть. Не появились лишь те контуры, в пределах которых листовые пластинки объединились так, что их можно было бы назвать листом. Первыми растениями, сделавшими этот шаг, являются голосеменные[3][4].
Папоротники размножаются спорами и вегетативно (вайями, корневищами, почками, афлебиями и так далее). Кроме этого, для папоротников характерно и половое размножение как часть их жизненного цикла.
Среди папоротников встречаются как травянистые, так и древесные формы жизни.
Лист папоротника — вайяТело папоротника состоит из листовых пластинок, черешка, видоизменённого побега и корней (вегетативного и придаточного). Листовидные органы папоротника называются вайями.
В лесах умеренной зоны обычно папоротники имеют короткий стебель, представляющий собой корневище, находящееся в почве. В стебле хорошо развита проводящая ткань, между пучками которой располагаются клетки основной — паренхимной ткани.
Вайи развёртываются над поверхностью почвы, вырастая из почек корневища. Эти органы обладают верхушечным ростом и могут достигать больших размеров; обычно они служат для выполнения двух функций — фотосинтеза и спорообразования. Спорангии размещаются на нижней поверхности вайи, в них развиваются гаплоидные споры.
Общий вид папоротникаВ жизненном цикле папоротника чередуется бесполое и половое поколение — спорофит и гаметофит. Преобладает фаза спорофита.
На нижней части листа раскрывается спорангий, споры оседают на земле, прорастает спора, появляется заросток с гаметами, происходит оплодотворение, появляется молодое растение.
У самых примитивных папоротников (ужовниковые) спорангии имеют многослойную стенку и не несут особых приспособлений для раскрывания. У более продвинутых — спорангий имеет однослойную стенку и приспособления к активному раскрыванию. Это приспособление имеет вид кольца. Уже среди примитивных папоротников прослеживается разноспоровость. У современных — небольшое число равноспоровых видов. Гаметофит равноспоровых обычно обоеполый. У примитивных он подземный и обязательно в симбиозе с грибами. У продвинутых гаметофиты надземные, зелёные и быстро созревающие. Они обычно имеют вид зелёной пластинки сердцевидной формы. Гаметофиты разноспоровых папоротников отличаются от равноспоровых (помимо своей раздельнополости) сильной редукцией, особенно мужского гаметофита. Женский гаметофит, потребляющий запасные питательные вещества у мегаспор, развит сильнее и имеет питательную ткань для будущего зародыша спорофита. При этом развитие таких гаметофитов происходит внутри оболочек мега- и микроспор.
По некоторым данным, папоротники произошли от плаунов, однако часть учёных считает, что хвощи, плауны, мхи и этот отдел произошли от псилофитов. В девонском периоде от споровых папоротников произошли семенные. Они относились к первым голосеменным растениям. От них произошли все остальные голосеменные и, вероятно, цветковые растения.
Для классификации папоротников в разное время были предложены схемы, и они зачастую плохо согласовывались друг с другом. Современные исследования поддерживают более ранние идеи, основанные на морфологических данных. В то же время, в 2006 году Аланом Смитом (англ. Alan R. Smith), ботаником-исследователем Калифорнийского университета в Беркли, и другими[5] была предложена новая классификация, основанная, в дополнение к морфологическим данным, на недавних молекулярных систематических исследованиях. Эта схема делит папоротники на четыре класса:
Последняя группа включает большинство растений, известных нам как папоротники.
Полная схема классификации, предложенная Смитом и другими в 2006 году, с учётом исправлений в части Cyatheaceae, предложенных в 2007 году группой Петры Корелл (англ. Petra Korall) и других[6]:
Классификация 2006/2007 года
Классификация 2014 года отличается преимущественно укрупнением семейств[7]:
Классификация 2014 года
- Подкласс Equisetidae Warm.
- Подкласс Ophioglossidae Klinge
- Подкласс Marattiidae Klinge
- Подкласс Polypodiidae Cronquist, Takht. & Zimmerm.
- Порядок Osmundales Link
- Порядок Hymenophyllales A.B.Frank
- Порядок Gleicheniales Schimp.
- Порядок Schizaeales Schimp.
- Порядок Salviniales Bartl.
- Порядок Cyatheales A.B.Frank
- Порядок Polypodiales Link
- Семейство Cystodiaceae J.R.Croft
- Семейство Lonchitidaceae C.Presl
- Семейство Lindsaeaceae C.Presl
- Семейство Saccolomataceae Doweld
- Семейство Dennstaedtiaceae Lotsy
- Семейство Pteridaceae E.D.M.Kirschn., подсемейства Cryptogrammoideae, Ceratopteridoideae, Pteridoideae, Cheilanthoideae, Vittarioideae
- Семейство Aspleniaceae Newman, подсемейства Cystopteridoideae, Rhachidosoroideae, Diplaziopsidoideae, Asplenioideae, Thelypteridoideae, Woodsioideae, Athyrioideae, Blechnoideae
- Семейство Polypodiaceae J.Presl & C.Presl, подсемейства Didymochlaenoideae, Hypodematioideae, Dryopteridoideae, Lomariopsidoideae, Tectarioideae, Oleandroideae, Davallioideae, Polypodioideae (трибы Loxogrammeae, Drynarieae, Platycerieae, Microsoreae, Polypodieae)
В 2016 году международной группой ботаников-систематиков (Pteridophyte Phylogeny Group) предложена консенсусная классификация ныне существующих таксонов[8]:
Классификация PPG I (2016)
Класс Lycopodiopsida Bartl.
корни, стебли, вайя папоротниковых растений
Папоротникообразные произрастают на планете Земля с незапамятных времен. Продолжительность их существования исчисляется миллионами лет. Растения имеют разнообразные жизненные формы, предпочитают влажные места обитания. Своеобразное строение папоротника делает его чрезвычайно приспособленным к выживанию.
Описание папоротниковидных растений
Что такое папоротник, есть разные версии. Ученые считают, что это прямые потомки древнейших растений – ринофитов. В процессе эволюции строение папоротникообразных стало более сложным, почему папоротники относят к высшим растениям, никого не удивляет. Это подтверждает:
- циклическое развитие растений;
- развитая система сосудов;
- приспособленность к наземной среде;

Папоротник в природе
По определению, папоротник – это многолетнее растение, относится к роду споровых растений. На вопрос о папоротнике: это кустарник или трава, можно ответить утвердительно в обоих случаях. Иногда еще бывает деревом.
Дополнительная информация. При описании папоротников нельзя не вспомнить их разнообразную цветовую гамму, доставляющую эстетическое удовольствие. Эти растения часто становятся настоящим украшением участков. Их несомненное достоинство – устойчивость к болезням и вредителям.
Важно! Как известно, папоротники не цветут, но в славянской мифологии цветок папоротника стал символом вечной любви и счастья. В ночь на Ивана Купала влюбленные тщетно ищут мифический цветок.
Биологическая классификация папоротниковых
Огромное количество видов папоротникообразных усложняет их классификацию. Такие попытки делались еще древними учеными. Предлагаемые схемы часто не согласовываются между собой. Основами классификации современных папоротников стали строение спорангия и некоторые морфологические признаки. Все разновидности делят на древние и современные.
К отделу папоротников относят следующие семь классов сосудистых растений, как вымерших, так и современных:
- Аневрофитопсиды (Aneurophytopsida) – древнейшая примитивная группа.
- Археоптеридопсиды (Archaeopteridopsida) – тоже древние представители, которые напоминали современные хвойные деревья.
- Кладоксилопсиды (Cladoxylopsida) – есть версии, что эта группа представляет слепую ветвь эволюции.
- Зигоптеридопсиды (Zygopteridopsida или Goenopteridopsida) – переходная группа к современным видам.
- Офиоглоссопсиды или ужовниковые (Ophioglossopsida) – современные папоротники.
- Мараттиопсиды (Marattiopsida) – многолетние растения мелких и крупных форм.
- Полиподиопсиды (Polypodiopsida – многолетние или реже однолетние растения разных размеров). Подразделяются на три подкласса: Polypodiidae, Marsileidae, Salviniidae.
История видов папоротниковых растений
История папоротникообразных началась в эпоху динозавров – 400 млн. лет назад. В благоприятном для них теплом и влажном климате тропических лесов папоротники господствовали на Земле. Некоторые виды достигали 30 м в высоту. Со временем климатические условия кардинально изменились. Сколько должно было произойти природных катаклизмов, чтобы исчезли такие исполины, как динозавры и древовидные папоротники.
При всем разнообразии современных папоротников они сильно отличаются от древнейших растений, уступая им в размерах и разнообразии форм. Но и сегодня это самая большая группа споровых – 300 родов и более 10 тыс. видов. Широкое распространение папоротникообразные получили, благодаря экологической пластичности и удивительным особенностям размножения.
Важно! Благоприятные для папоротников климатические условия в наши дни сохранились в тропиках и субтропиках, где древовидные папоротники достигают 20 м.
Распространенность в природе, примеры видов
Когда и где растет папоротник, зависит от тепла и влажности местности. Местом обитания вездесущих растений могут быть:
- нижний и верхний ярусы лесов;
- болота, реки и озера;
- овраги и сырые луга;
- расселины скал;
- стены домов;
- обочины дорог.

Страусник
В умеренных широтах можно встретить сотни травянистых представителей папоротников. Краткий обзор некоторых видов:
- Орляк обыкновенный. Легко узнаваем по листьям в форме раскрытого зонтика. Распространен в сосновых лесах, подходит для употребления в пищу.
- Щитовник мужской. Травянистое растение с листьями до 1,5 м в длину, встречается очень редко. Экстракт Щитовника используется в медицине как антигельминтное средство.
- Кочедыжник женский. Крупное растение с листьями изящной формы.
- Страусник обыкновенный. Крупный красивый папоротник. За счет длинных корневищ образует целые заросли. Используется в озеленении участков. Вареные листья Страусника съедобны.
- Осмунда. Растение с коротким корневищем и длинными блестящими листьями произрастает в Восточной Азии и Северной Америке.
- Многорядник. Его листья темно-зеленого цвета, расположены рядами.
Важно! Орляк настолько быстро растет, что его посадки следует ограничивать различными ограждениями, вкопанными в землю.
Для домашнего выращивания подходят:
- Костенец;
- Нефролепис;
- Даваллия;
- Асплениум;
- Диксония;
- Птерис.
Эти растения замечательно украшают домашний интерьер. Очень красив сорт Юниор с гофрированными листьями.

Домашнее растение
Интересные факты:
- Хецистортерис пумила и Азолла карилиниана – самые низкие растения. Их длина не превышает 12 мм.
- Эпифиты прекрасно себя чувствуют на деревьях и лианах.
- В горных районах можно встретить Венерин волос – удивительное растение с красивыми ажурными листьями.
- Древовидные стволы крупных папоротников в тропиках используют в качестве строительного материала.
- Marsilea quadrifolia прекрасно чувствует себя под водой.
- Дикраноптерис (Dicranopteris) имеет листовые черешки металлической прочности.
Редкие виды папоротникообразных:
- Щитовник гребенчатый;
- Фегоптерис связывающий;
- Многорядник Брауна;
- Асплений стенной;
- Гроздовник многораздельный.
- Папоротник Сальвиния плавающая занесен в Красную книгу Беларуси.
Важно! Растения с красивыми листьями имеют декоративное значение, часто используются в ландшафтном дизайне и при составлении флористических композиций.
Сколько живут папоротники
Вопрос «сколько лет живет папоротник» непростой. Продолжительность жизни зависит от места его произрастания и вида. В умеренных широтах наземная часть папоротника отмирает с наступлением холодов, в тропиках может расти несколько лет. Придаточные корни каждые 4 года заменяются новыми, непосредственно корневище сохраняет жизнеспособность до 100 лет. Эта особенность дает возможность растению выживать в любых условиях.
Особенности питания растения
Папоротникообразные питаются, добывая необходимые питательные вещества при помощи корней и листьев. Необходимые микроэлементы и воду растение всасывает из почвы. Листья участвуют в процессе фотосинтеза, преобразуя углекислый газ в органические кислоты. Так папоротник получает крахмал и сахар, нужные для жизнедеятельности всех органов.
Разбор строения папоротниковых растений
Предки папоротниковых растений имели примитивное строение. В процессе эволюции оно стало более сложным.
Стебли
Стебель папоротников малоразвит, небольших размеров. Его называют корневищем. Конечно, тропические папоротники с одеревеневшими стволами являются исключением. Вьющееся корневище может простилаться на большие расстояния.
Листья, вайи
Лист папоротника гораздо массивнее стебля. Они не совсем обычные, имеют отличительные особенности строения и роста, разнообразные формы. Чаще они рассеченные, перистые. Вайи – так называются листья у папоротника. Черешок листа прикреплен к подземной части стебля – корню или корневищу. Рассматривая их, трудно понять, где заканчивается стебель, и на каком уровне начинается лист. Интересная особенность вайи – рост верхушкой, которая представляет собой свернутый и постепенно разворачивающийся завиток в виде улитки.

Вайи
Развитие листьев начинается в почках под землей и длится до двух лет. Только на третий год они могут появиться над землей. Вследствие верхушечного роста вайи папоротников достигают очень крупных размеров.
У большинства растений вайи участвуют в процессе фотосинтеза, вегетации и одновременно в образовании спор. Споры появляются в сорусах, которые расположены на нижней стороне листьев в виде одиночных или групповых бугорков.
Корневая система
Корневая система состоит из мощного корневища и многочисленных придаточных корней. Проводящая ткань на стебле и корнях всасывает воду и по сосудистым пучкам продвигает ее к листьям.
Органы размножения
Жизнь папоротника делится на два цикла: длинный бесполовой – спорофит и короткий половой – гаметофит. Органы размножения папоротника – спорангии, где находятся споры, расположены на нижней части листьев. Созревшие споры высыпаются из лопнувшей спорангии и разносятся ветром далеко от материнского растения. Спорообразование некоторые ученые сравнивают с цветением других растений.
Из огромного количества спор выживает только часть. Половая фаза начинается, когда спора прорастает при благоприятных условиях в гаплоидный заросток (гаметофит), который похож на зеленую пластину в виде сердца размером в несколько мм. На нижней стороне заростка формируются женские и мужские половые органы – антеридии и архегонии. Образовавшиеся в них яйцеклетки и сперматозоиды во влажную погоду сливаются, и образуется зигота, из которой развивается зародыш молодого растения – спорофит.
Растение может также размножаться вегетативным способом, когда на стеблях и корнях образуются выводковые почки. Это важно для любителей-садоводов, выращивающих некоторые виды на своих участках.
Важно! Замечено, что красивые редкие виды размножаются только спорами.
Сравнение с другими травянистыми растениями
Папоротниковидные – это папоротники, хвощи и плауны. Все они размножаются спорами и имеют общее происхождение.
У папоротников есть характерные отличительные признаки от других травянистых растений:
- От водорослей они отличается корневищем и сложными листьями.
- У мхов и папоротников происходит чередование поколений гаметофита и спорофита. У мхов преобладает гаметофит, у папоротник – спорофит. Наличие проводящей ткани в виде сосудистых пучков делает представителей папоротниковидных растений более приспособленными к наземному образу жизни.
- В отличие от цветковых растений, размножаются спорами и не цветут.
Благодаря богатому химическому составу, папоротник имеет полезные для людей свойства. Используется в лекарственных целях и в кулинарии, подходит для засолки. Народная медицина тоже не обошла своим вниманием чудесные растения.
Папоротниковые — Википедия
Материал из Википедии — свободной энциклопедии
Па́поротниковые, или Настоя́щие па́поротники, или Многоножковидные, или Полиподиопсиды (лат. Polypodiópsida) — класс папоротниковидных растений.
Папоротниковые — гораздо более молодой и более крупный класс папоротников, нежели Мараттиевые — представители другого класса папоротниковидных. У всех настоящих папоротников листья-вайи несут спорангии со стенкой толщиной в одну клетку, и поэтому их называют тонкоспорангиатными. Большинство настоящих папоротников имеет специализированные структуры, способствующие раскрыванию спорангиев — так называемое кольцо, или отдельные группы толстостенных клеток. При высыхании спорангия тонкостенные клетки сжимаются быстрее, нежели стенки кольца, и благодаря этому стенка спорангия разрывается, освобождая споры.[2]
В варианте, предложенном Аланом Р. Смитом (Калифорнийский университет в Беркли) с соавторами[3]. Новая классификация основывается на филогенетических исследованиях, проведённых начиная с 1994 года.
- Порядок Осмундовые с единственным семейством того же названия — самая примитивная группа среди современных папоротников. Осмундовые — одна из древнейших групп настоящих папоротников — остатки растений, близких к этому семейству, встречаются уже в отложениях конца каменноугольного периода. Временем их расцвета был мезозой, а до наших дней дожило не так много представителей группы. Многие осмундовые имеют очень крупные листья, размер которых у осмунды королевской (Osmunda regalis L.) достигает двух метров. На листьях расположены не собранные в группы спорангии, не имеющие кольца (его роль играют две группы толстостенных клеток)[2].
- Гименофилловые папоротники иногда называют «плёнчатыми папоротниками», чем подчёркивается строение их тонких, прозрачных листьев, ткани которых подверглись значительной редукции в результате жизни во влажной среде. Гименофилловые приспособились к обитанию в очень тёмных и очень влажных местообитаниях — в нижних ярусах тропического леса, по берегам горных рек, на стволах околоводных деревьев, на камнях водопадов и т. п. В связи с приспособлением к подобным местообитаниям возникла специализация, которая привела к редукции как спорофита, так и гаметофита гименофилловых. Так, например, листья гименофилловых лишены устьиц и абсолютно проницаемы для воды. Поскольку такие листья так же легко теряют воду, как и её поглощают, Гименофилловые развили вторичную пойкилогидридность, позволяющую им переносить засушливые периоды без серьёзного вреда для своих клеток[2].
- Папоротники из семейств Матониевые и Диптерисовые являются «живыми ископаемыми» — остатками когда-то очень богатой меловой флоры тропического пояса. Они сохранились отдельными островами на вершинах гор Индонезии и Новой Гвинеи, зачастую на расстоянии нескольких сотен километров друг от друга[2].
- Листья схизейных, как правило, разделены на спороносную и вегетативную части, причем спороносная часть может нисколько не напоминать лист, а быть похожей на палочки, спирали, кисточки и тому подобные структуры. Род Лигодиум (Lygodium Swartz) этого семейства отличается гигантскими листьями: их длина может достигать тридцати метров. Они способны к верхушечному росту (как и у большинства настоящих папоротников), что легко объясняет их возникновение[2].
- К порядку Сальвиниевые — водных папоротников — относится ряд приспособившихся к обитанию в воде и переувлажнённых местообитаниях папоротников, развивших разноспоровость. Развитие спор и строение заростков этих папоротников напоминает Selaginella — Селагинелла, или Плаунок, хотя никаких родственных отношений между этими группами нет. Таким образом, независимое возникновение разноспоровости у плауновидных и у настоящих папоротников представляет собой пример настоящей конвергенции. Наиболее распространенные роды сальвиниевых — Марсилия и Сальвиния. Первая представляет собой небольшие прибрежные и водные растеньица, больше всего напоминающие четырёхлистный клевер. Спорангии марсилии собраны под общей оболочкой в так называемый спорокарп, который обладает весьма сложными механизмами раскрывания. Сальвинии — это плавающие на поверхности воды ряскообразные (но значительно более крупные) растения с очень интересным диморфизмом листьев — два из трёх листьев в мутовке плавающие, фотосинтезирующие, а третий — корневидный, обеспечивающий устойчивость растения и всасывание воды. Кроме того, на нижней поверхности обычных листьев сальвинии развивается киль, а на верхней — блестящие водоотталкивающие клетки[2].
- Представители порядка Циатейные — так называемые древовидные папоротники, обитающие, как правило, в высокогорных туманных лесах, где испарение воды стеблем незначительно. Пробка у них не развивается, и это сильно ограничивает распространение циатейных. Вторичного утолщения коры представители этого порядка, в отличие от настоящих деревьев, не имеют, и древесина их состоит лишь из первичных тканей — проводящих пучков и отмерших клеток паренхимы[2].
- Порядок Polypodiales — Многоножковые
- Семейство Lindsaeaceae (= Cystodiaceae, Lonchitidaceae)
- Семейство Saccolomataceae
- Семейство Dennstaedtiaceae Pic. Serm. — Деннштедтиевые (= Hypolepidaceae, Monachosoraceae, Pteridiaceae)
- Семейство Pteridaceae (= Acrostichaceae, Actiniopteridaceae, Adiantaceae, Anopteraceae, Antrophyaceae, Ceratopteridaceae, Cheilanthaceae, Cryptogrammaceae, Hemionitidaceae, Negripteridaceae, Parkeriaceae, Platyzomataceae, Sinopteridaceae, Taenitidaceae, Vittariaceae)
- Семейство Aspleniaceae A.B. Frank — Костенцовые
- Семейство Thelypteridaceae Pic. Serm. — Телиптерисовые
- Семейство Woodsiaceae — Вудсиевые (= Athyriaceae, Cystopteridaceae)
- Семейство Blechnaceae (= Stenochlaenaceae)
- Семейство Onocleaceae — Оноклеевые
- Семейство Dryopteridaceae Herter — Щитовниковые (= Bolbitidaceae, Elaphoglossaceae, Hypodematiaceae, Peranemataceae)
- Семейство Lomariopsidaceae (= Nephrolepidaceae)
- Семейство Tectariaceae
- Семейство Oleandraceae
- Семейство Davalliaceae
- Семейство Polypodiaceae — Многоножковые (= Drynariaceae, Grammitidaceae, Gymnogrammitidaceae, Loxogrammaceae, Platyceriaceae, Pleurisoriopsidaceae)
- Порядок Многоножковые наиболее богат видами среди всего класса. К одноимённому семейству Многоножковые принадлежит, например, Многоножка, широко распространённая в горах Европы. Спорангии у многоножки собраны в округлые скопления — сорусы, а корневище покрыто многочисленными рыжими волосками, предохраняющими от излишнего испарения. Другой представитель семейства — Платицериум, или Олений рог, знаменит способностью образовывать себе ёмкость для питательного субстрата. Этот эпифитный папоротник имеет два типа листьев: обычные (вильчато рассечённые) и щитковидные, образующие «чехол» вокруг основания растения. Со временем внутри чехла накапливается перегной, и папоротник оказывается в «горшке», достигающем у некоторых экземпляров массы в 100 кг[2].
- Smith, A. R., K. M. Pryer, et al. A classification for extant ferns (англ.) // Taxon. — 2006. — Vol. 55(3). — P. 705—731.
- Rothwell, G. W. and K. C. Nixon. How does the inclusion of fossil data change our conclusions about the phylogenetic history of euphyllophytes // Int. J. Plant Sci.. — 2006. — Т. 167(3). — С. 737—749.
- Kramer, K. U. Notes on the Higher Level Classification of the Recent Ferns // K. Kubitzki, K. U. Kramer and P. S. Green The Families and Genera of Vascular Plants: Pteridophytes and Gymnosperms. — New York: Springer-Verlag, 1990. — Т. 1. — С. 49—52.
- Charles T. Druery. British ferns and their varieties. — London: E. P. Dutton and Co., 1912?.